V. LA ENERG�A CELULAR
LOS CAP�TULOS anteriores nos dieron una visi�n de la membrana Como estructura de envoltura y protecci�n para la c�lula que es al mismo tiempo activa y capaz de realizar el intercambio de materiales con el exterior. Tambi�n ya se hizo menci�n al hecho de que hay otras membranas y estructuras formadas a base de ellas que tienen funciones especiales dentro de las c�lulas, y en el cap�tulo anterior se se�alaron las relaciones que existen entre el transporte y la energ�a.
Uno de los problemas estrechamente ligado con esta relaci�n entre el transporte y la energ�a es la necesidad que tienen los organismos de convertir la energ�a de las formas en que se encuentra, ya sea directa, como la luminosa o la qu�mica de los enlaces entre los �tomos de los az�cares y otras sustancias, en otras aceptables para los diferentes sistemas que la c�lula requiere para funcionar.
Hay m�quinas y veh�culos que pueden funcionar con agua y le�a o carb�n, porque cuentan con una caldera y un sistema capaz de aprovechar la presi�n del vapor de agua para producir trabajo mec�nico, y otros que deben utilizar combustibles m�s finos como la gasolina, por ejemplo. Una m�quina de vapor no funciona inyect�ndole gasolina en los pistones, ni un motor de gasolina con carb�n como combustible. Ello se debe a que estas m�quinas se han construido de manera que no es siquiera posible utilizar indistintamente los combustibles en las diferentes m�quinas. Algo semejante sucede con las c�lulas, y en general con los seres vivos. No obstante que pr�cticamente toda la energ�a que se utiliza para mantener la vida en la tierra proviene del sol, es muy claro que ning�n ser vivo puede realizar directamente sus funciones con la energ�a luminosa. Igualmente, aunque la glucosa es uno de los combustibles por excelencia para los seres vivos, tampoco es posible hacer que una fibra muscular se contraiga directamente agregando este az�car. Fue entonces necesario que los seres vivos desarrollaran sistemas adecuados para proveerse de los combustibles que s� pueden ser utilizados por las diferentes piezas de la maquinaria celular.
El combustible celular m�s importante, que ya se mencion� en el cap�tulo anterior, es el ATP, mol�cula que contiene tres fosfatos unidos uno a otro, y que as� como libera energ�a al romperse los enlaces entre sus fosfatos, tambi�n se puede formar por la uni�n del ADP que s�lo contiene dos fosfatos y un fosfato m�s, adem�s de una cantidad importante de energ�a. Ya qued� claro tambi�n que el ATP se puede emplear directamente para que algunos sistemas de transporte muevan iones activamente a trav�s de las membranas; esta sustancia tambi�n puede proporcionar directamente la energ�a necesaria para que las fibras musculares se contraigan y produzcan as� el movimiento. �stos son s�lo dos de los ejemplos de la utilidad de esta sustancia como "combustible" celular, es decir, como la sustancia que s� puede ser aprovechada directamente por los sistemas que realizan diferentes tipos de trabajo en las c�lulas.
Visto as� el problema, es f�cil imaginar que la naturaleza haya desarrollado y perfeccionado diferentes tipos de mecanismos para sintetizar tan importante mol�cula a partir de sus componentes y proveer a las c�lulas el combustible que requieren constantemente para realizar sus funciones.
Aunque hay otros compuestos que podr�amos considerar como combustibles para las c�lulas, es el ATP el que probablemente tenga la funci�n m�s universal en cuanto a los organismos y sistemas que lo aprovechan para funcionar.
LA LUZ, LA ENERG�A Y EL ALIMENTO
Dado que la energ�a que mantiene la vida sobre la tierra es la que proviene del sol, es de primordial importancia la funci�n de capturarla y convertirla en formas directamente aprovechables por los seres vivos, como el ATP, o inclusive almacenarla en otros compuestos, como los az�cares. A este proceso se le llama fotos�ntesis.
Hay numerosas variantes en la naturaleza en cuanto a los sistemas que se encargan
de transformar la energ�a luminosa, generalmente del sol, en forma directamente
aprovechables por los seres vivos y que permiten a las c�lulas producir ATP
y otras sustancias; sin embargo, el esquema es m�s o menos constante y se le
puede describir de una manea general, a sabiendas de que no es exactamente igual
en todos los casos. Este tema es central al tratar sobre las membranas, debido
a que, para que el sistema funcione, requiere realizarse en una estructura membranosa
cerrada y compleja: el cloroplasto. En la Figura 31 se muestra un esquema de
la estructura de este organelo. El cloroplasto es una estructura membranosa
cerrada, y tanto la existencia de membranas impermeables, como la de estructuras
cerradas, son requisitos indispensables para que se realice la fotos�ntesis.
Las estructuras membranosas cerradas, llamadas tilacoides, son peque�as y aplastadas,
se apilan unas sobre otras seg�n se muestra en la figura, y est�n rodeadas tambi�n
por otras dos membranas. Pero tal vez lo importante de se�alarlo es que los
procesos principales de la fotos�ntesis tienen lugar en los tilacoides, y adem�s
es necesaria su integridad para que su proceso se lleve a cabo.
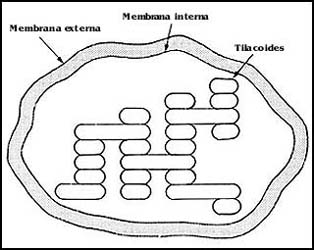
Figura 31. Esquema de la estructura de un cloropasto. La estructura completa est� rodeada por una doble membrana, en cuyo interior se encuentran los tilacoides. Los tilacoides son formaciones membranosas cerradas en forma de discos apilados, en los cuales se llevan a cabo los procesos de aprovechamiento de la luz.
Para capturar la luz, adem�s de la clorofila, los cloroplastos y otras estructuras encargadas de realizar la fotos�ntesis utilizan una serie de pigmentos, es decir sustancias coloreadas. Dicho sea de paso, las sustancias que tienen color deben esta propiedad al hecho de que absorben ciertos tipos de luz y dejan pasar otros. Dado que la fotos�ntesis consiste en la conversi�n de energ�a luminosa en energ�a qu�mica, los cloroplastos requieren antes que nada un sistema eficiente para capturar la luz. Esto se logra en primer lugar mediante la existencia de un gran n�mero de estos organelos en las hojas de las plantas, pero gracias tambi�n a la existencia de diferentes sustancias de color llamadas pigmentos antenas, y no s�lo, como es creencia com�n, por la existencia de la clorofila. Estos pigmentos simplemente sirven para recoger la mayor cantidad de luz posible y aumentar la eficiencia del proceso, evitando que algunos rayos de luz atraviesen el cloroplasto y se disipen en el medio o sean interceptados por compuestos que no tengan nada que ver con la fotos�ntesis. La luz capturada simplemente pasa de unos a otros de los pigmentos hacia la parte que efectivamente tiene la capacidad de utilizarla, que recibe el nombre de centro de reacci�n (Figura 32).
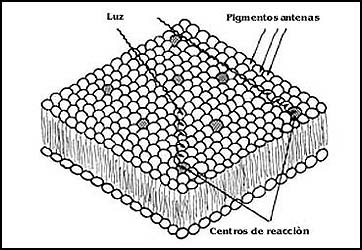
Figura 32. Los centros de reacci�n y los pigmentos antenas en la membrana del tilacoide. Los pigmentos antenas aumentan la eficiencia en la captura de la luz y la transmiten a los centros de reacci�n.
Este centro de reacci�n es una estructura complicada cuyo funcionamiento no se conoce con detalle, pero que utilizando la energ�a de la luz previamente capturada por los pigmentos antenas lo primero que hace es tomar un par de electrones que provienen del agua, produciendo su descomposici�n y liberando ox�geno (Figura 33 ). El proceso recibe el nombre de fot�lisis y es de extraordinaria importancia, pues es la que provee de ox�geno a los seres vivos que, como los humanos, dependemos estrictamente de su presencia en el ambiente.
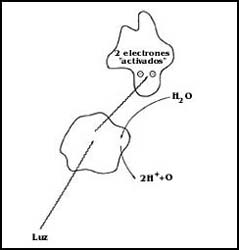
Figura 33. La fot�lisis del agua. Utilizando la energ�a de la luz, se libera oxígeno y los hidr�genos se descomponen para dar dos protones y dos electrones. Estos se "activan" o energizan tambi�n con la energ�a de la luz.
El par de electrones que se toma de la mol�cula de agua, utilizando la misma energ�a del sol, se "activa" o cambia su energ�a mediante la transferencia a un compuesto, como se muestra en la Figura 34. De este compuesto, los electrones pueden luego ser transferidos sucesivamente a otros en una especie de cadena, muy semejante a la cadena respiratoria. Lo importante de ello es que esta transferencia sucesiva de los electrones activados de unas sustancias a otras, como la cadena respiratoria, es un proceso que produce energ�a, la cual no se desperdicia; se aprovecha en la transferencia misma para, simult�neamente, bombear protones (H+) al interior del tilacoide, generando as� una diferencia de concentraci�n de �stos, que, como ya se se�al� anteriormente (cap�tulo IV), equivale a otra forma de energ�a en la que se ha convertido la energ�a luminosa.
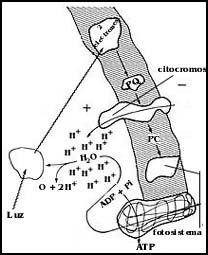
Figura 34. El funcionamiento del fotosistema II. Los electrones "activados" con la energ�a de la luz, caen a trav�s de varios intermediarios hacia el fotosistema I. En la "ca�da" de los electrones, la energ�a se aprovecha para bombear protones (H+) hacia el interior del tilacoide.
Los protones que se han acumulado as� dentro de la estructura membranosa tienden a salir, pero la membrana les impide el paso; �ste s�lo es posible a trav�s de una enzima colocada en ella, que tiene la capacidad de aprovechar la energ�a con que los H+ tienden a regresar al exterior del tilacoide. La energ�a es suficiente y se utiliza para sintetizar el ATP a partir de sus componentes, ADP, y P (fosfato).
Esta enzima que sintetiza el ATP, y a la que se le ha llamado ATP sintetasa, es interesante, pues en realidad es capaz de formar ATP de sus componentes, el ADP y el fosfato; puede funcionar como una de las "bombas" que se describieron en el cap�tulo IV, rompiendo el ATP y utilizando la energ�a para bombear protones al interior del tilacoide, pero cuando hay luz empieza a funcionar al rev�s, es decir, la presi�n de los protones que se han bombeado al interior la obliga ahora a sintetizar ATP a partir de ADP y fosfato, utilizando para ello la energ�a que se ha acumulado en forma de una diferencia de concentraci�n de hidrogeniones en ambos lados de la membrana.
Esto es lo que sucede en el fotosistema II del cloroplasto; sin embargo, el
fotosistema I puede tomar los mismos electrones que han llegado al final de
la cadena de electrones del fotosistema II y activarlos nuevamente, utilizando
tambi�n para ello la energ�a luminosa. De nuevo los electrones activados pueden
ser transportados por varios compuestos para llegar finalmente a producir otra
sustancia llamada NADPH (nicot�n aden�n dinucle�tido fosfato, reducido),
que luego es �til para la s�ntesis de la glucosa, pues proporciona de cierta
manera los hidr�genos que se requieren para la s�ntesis de la mol�cula de glucosa
a partir de C02, y es �sta el producto final
de la fotos�ntesis (Figura 35).
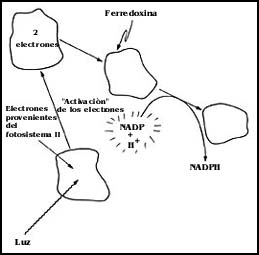
Figura 35. El fotosistema I. Los electrones que provienen del fotosistema II son activados de nuevo con energ�a de la luz. Son entregados finalmente al NADP para producir NADPH.
Una vez que el cloroplasto ha producido estos compuestos, el ATP y el NADPH, el resto del proceso se reduce a una serie m�s o menos complicada de pasos enzim�ticos, que tambi�n tienen lugar en el cloroplasto fuera de los tilacoides, y despu�s de los cuales el balance para sintetizar una mol�cula de glucosa ser�a el siguiente:
6CO2 + 18ATP
+ 12 NADPH + 12H2O ®
C6H12O6
+ 18 P + 12NADP
Como ya se ha mencionado, la glucosa puede considerarse como una de las formas de almacenamiento de la energ�a luminosa del sol por parte de las plantas. El resumen del proceso es como sigue:
1) La energ�a luminosa se convierte, en parte, en un potencial el�ctrico en ambos lados de la membrana, y en parte en NADPH, que representa la manera como se agregan �tomos de hidr�geno a las mol�culas durante su s�ntesis.
2) Parte de la energ�a que se convirti� primero en la diferencia de potencial el�ctrico, se convierte a su vez en una forma de energ�a qu�mica que es la de los enlaces entre el ADP y el fosfato: el ATP.
3) Finalmente, tanto los hidr�genos del NADPH como la energ�a
del ATP son utilizados para unir una por una las mol�culas de C02
y formar mol�culas de glucosa.
La glucosa no representa realmente la forma en la que se "almacena" la energ�a estrictamente hablando; la mayor�a de las c�lulas y los organismos vegetales suelen almacenar pol�meros de ella, de los cuales el m�s conocido es el almid�n, que las plantas guardan en las semillas, tub�rculos y otros materiales, y que muchos de los animales utilizamos para alimentarnos y recuperar de �l la energ�a almacenada.
Adem�s, las c�lulas mismas encargadas de la fotos�ntesis tambi�n tienen otras formas de aprovechar el mismo ATP producido durante la fotos�ntesis en numerosos procesos adicionales que requieren energ�a, como es el caso de cualquier otra c�lula.
Es un hecho conocido que la energ�a y nuestra respiraci�n est�n relacionadas entre s�. Todos sabemos que si corremos o hacemos alg�n tipo de ejercicio f�sico aumenta la frecuencia y la intensidad de nuestros movimientos respiratorios, que de alguna forma sirven para proporcionar a nuestros tejidos una mayor cantidad de ox�geno. Sin embargo, fuera de esto es poco conocido el significado o la base real de la relaci�n entre la respiraci�n, o sea el consumo de ox�geno, y la conversi�n o aprovechamiento de la energ�a por nuestras c�lulas y tejidos. Pero todo parte de una ecuaci�n muy sencilla, la de la oxidaci�n de la glucosa, y que es la siguiente:
C6H12O6 + 6O2 ® 6H2O + energ�a
Glucosa + oxígeno ® bi�xido de carbono + agua + energ�a
De paso, �sta es la ecuaci�n que representa tambi�n la combusti�n de la glucosa si se la quema en presencia de ox�geno, como si fuera cualquier otro material combustible. S�lo que si se quema este az�car en presencia del aire (ox�geno), la energ�a se desprende en forma de calor y pasa sin utilizarse al medio ambiente. Los seres vivos tienen la capacidad de realizar el mismo proceso, pero en lugar de desperdiciar la energ�a como calor la pueden aprovechar, transform�ndola nuevamente en la energ�a de los enlaces del ATP, la forma universal del combustible celular.
De lo anterior, y de lo que ya se ha revisado, es claro que se establece un ciclo entre las plantas y los animales; las primeras liberan ox�geno en la fotos�ntesis, y los segundos lo toman para respirar, como se muestra en el esquema de la Figura 36. Adem�s, si las plantas producen glucosa (o almid�n) y los animales la utilizan, tambi�n hay un ciclo en este sentido, y es as� como el sol es en �ltima instancia la fuente de la energ�a para todos los seres vivos.
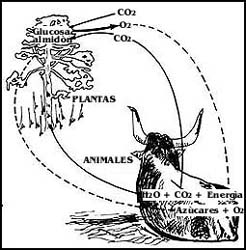
Figura 36. El ciclo de materiales y energ�a entre plantas y animales. Los animales aprovechan mol�culas m�s o menos grandes y la energ�a acumulada en ella y el ox�geno que las plantas producen. Las plantas aprovechan el CO2 que producen los animales.
Pero la obtenci�n de la energ�a que las plantas han almacenado en forma de glucosa no es un proceso sencillo; para el efecto, o hay un sistema que opera en las bacterias o se utiliza uno que tiene lugar en un organelo intracelular, dise�ado principalmente con esta funci�n. Este organelo es la mitocondria, y la funci�n recibe el nombre de fosforilaci�n oxidativa.
La mitocondria es una estructura membranosa cerrada; de hecho est� formada por una doble membrana: la interna est� plegada sobre s� misma, formando unas estructuras llamadas crestas, dentro de las cuales hay un espacio llamado matriz, que contiene numerosas enzimas y otras sustancias (Figura 2). Adem�s, debemos insistir en que para que se realice la fosforilaci�n oxidativa es necesario que exista la integridad de la membrana interna y su impermeabilidad. En las bacterias, el proceso se lleva a cabo en la membrana que las rodea.
La fosforilaci�n oxidativa funciona de una manera semejante al fotosistema II de la fotos�ntesis; tambi�n tiene una cadena de transporte de hidr�genos y electrones, pero en ella su destino final es el ox�geno, y aqu�llos no necesitan ser activados, pues tienen ya un estado "energizado", de tal forma que en su camino hacia el ox�geno se libera energ�a. Es necesario se�alar que la energ�a de estos electrones proviene de los enlaces de la mol�cula de glucosa; esa energ�a a su vez tiene su origen en la del sol, y fue capturada durante la fotos�ntesis. Los electrones provienen de diversas sustancias, que a su vez resultan de la degradaci�n de la glucosa; el mecanismo de producci�n de estas sustancias es el siguiente:
1) Despu�s de un largo proceso previo, de una mol�cula de glucosa resultan
dos de acetato, que es un fragmento de dos �tomos de carbono, como se muestra
en la Figura 37. Este fragmento de dos �tomos de carbono entra a un proceso
c�clico en el cual se degrada, pero para ello debe unirse a uno de los componentes
de ese ciclo. En el esquema de la figura se puede ver que en dos de los pasos
del ciclo metab�lico salen sendas mol�culas de CO2,
que representan los dos �tomos de carbono del acetato. Pero la otra cosa importante
es que en este ciclo, llamado ciclo de los �cidos tricarbox�licos o ciclo de
Krebs, en honor de su ilustre descubridor, Hans Krebs, una serie de sustancias
cede hidr�genos por pares (2H) a otra, que es el NAD, convirti�ndolo
en lo que se conoce como NADH. En cierta forma, el proceso equivale
a tener �tomos de hidr�geno que luego se pueden unir con el ox�geno durante
la respiraci�n para formar agua.
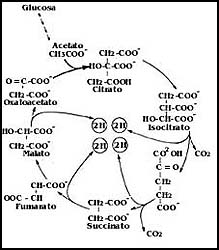
Figura 37. El ciclo de los �cidos tricarbox�licos: entra una mol�cula de acetato (dos �tomos de C) salen dos mol�culas de CO2 y cuatro pares de hidr�genos.
2) A continuaci�n, y gracias a la tendencia que tiene el hidr�geno para unirse con el ox�geno, de todos conocida, los pares de �ste que se han producido en el ciclo de los �cidos tricarbox�licos son transferidos hasta el ox�geno, pero no en un solo paso, sino a trav�s de una serie sucesiva de compuestos que se encuentran localizados en la membrana interna de la mitocondria. Tal parece que esta transferencia sucesiva est� dise�ada para que el proceso sea gradual y se pueda aprovechar al m�ximo la energ�a de esos electrones en su paso hacia el ox�geno. Finalmente se produce una mol�cula de agua.
Al sistema de transferencia de los hidr�genos (o electrones) se le conoce como cadena de transporte de electrones o cadena respiratoria. El segundo nombre es f�cil de entender; el otro se refiere al hecho de que, estando el �tomo de hidr�geno compuesto de un prot�n y un electr�n solamente, hay componentes de la cadena respiratoria que lo pueden transportar completo (prot�n y electr�n), y otras en las que s�lo se transportan los electrones.
3) Pero una de las caracter�sticas importantes de este transporte de electrones es que la energ�a de su transferencia representa realmente una "ca�da" de nivel, y, de la misma manera que en la ca�da de un cuerpo o de una cantidad de agua, se libera energ�a en el proceso. En el caso de los electrones en la cadena respiratoria, la energ�a no se desperdicia, pues simult�neamente a la ca�da de aqu�llos, los componentes tienen la capacidad de utilizar la energ�a para expulsar hidrogeniones (H+) de la mitocondria para crear una diferencia en su concentraci�n que, como ya mencionamos, tiene dos elementos de energ�a, la diferencia de concentraci�n y la del n�mero de cargas en ambos lados de la membrana. Es as� que esencialmente, y en un primer paso, la energ�a de los electrones al caer hacia el ox�geno, y que proviene originalmente de los enlaces de la glucosa o de otras sustancias, se transforma en un potencial electroqu�mico dado por la diferencia de concentraci�n de los hidrogeniones.
4) As�, lo �nico que se requiere ahora es un mecanismo capaz de aprovechar
esta energ�a representada por el potencial de la membrana para la s�ntesis del
ATP. La mitocondria, de la misma forma que el cloroplasto, tiene
una enzima en la membrana, otra ATP sintetasa, que puede aprovechar
la energ�a con que los hidrogeniones tienden a regresar a la mitocondria, para,
al paso de ellos, sintetizar ATP a partir de ADP y
fosfato, como se muestra en la Figura 8.
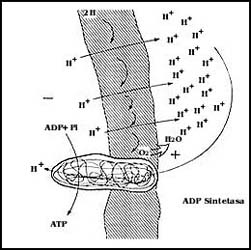
Figura 38. La s�ntesis del ATP en las mitocondrias. La energ�a representada por la acumulaci�n de H+ en el exterior es aprovechada por la ATP sintetasa para unir al ADP y al fosfato (P) para sintetizar ATP.
SEMEJANZAS DE LOS SISTEMAS DE TRANSPORTE Y LOS DE TRANSFORMACI�N DE
ENERG�A
Estos dos sistemas, fotos�ntesis y fosforilaci�n oxidativa, proporcionan a las c�lulas la mayor parte de la energ�a que necesitan para funcionar. De alguna manera, los mecanismos involucrados en ambos procesos no difieren gran cosa de los empleados por los sistemas de transporte. Las cadenas de transporte de electrones, tanto del fotosistema II de la fotos�ntesis, como la de la mitocondria, pueden verse simplemente como sistemas de bombeo de H+ o translocadores primarios, que adem�s tienen funciones muy bien definidas para transportar iones y otros metabolitos. De hecho, tanto el potencial negativo interno, como la diferencia de concentraci�n de H+ en ambos lados de la membrana, sirven tambi�n para que la mitocondria tome otros elementos del medio en que se encuentra requeridos para realizar sus funciones. �stas son tal vez de las m�s complicadas realizadas por la membrana mitocondrial que existen entre los seres vivos. Esta membrana es una de las que muestran un mayor contenido de prote�nas, y esto se debe seguramente a la gran cantidad de funciones que tiene que desempe�ar, y de las cuales s�lo hemos mencionado aqu� algunas.
Estos procesos, tanto la fosforilaci�n oxidativa, como la fotos�ntesis, tambi�n pueden realizarse en las bacterias, pero en ellas no existe m�s que la membrana externa. Tanto la fotos�ntesis como la fosforilaci�n oxidativa se realizan con variaciones en las bacterias, sobre todo en el hecho de que los movimientos de protones en un proceso y otro tienen que realizarse con el exterior. Inclusive, la semejanza de estos procesos entre las bacterias y los cloroplastos y mitocondrias, as� como otros hechos, hacen suponer que realmente esos organelos surgieron en el curso de la evoluci�n, porque dentro de alguna c�lula que ya exist�a, se incluyeron bacterias que les permitieron desarrollar funciones adicionales y mejorar sus sistemas de conversi�n de energ�a.