IV. CORRE, QUE TE ALCANZO. INTERACCIONES DEPREDADOR-PRESA
| Un gavil�n se levanta un pich�n que est� dormido todo el palomar se espanta y queda despavorido. Son de la Tierra Caliente del Balsas |
LA SEGUNDA gran clase de interacci�n entre dos especies es la
relaci�n llamada depredador-presa. Corresponde a los cambios demogr�ficos representados
por los signos +-, lo que significa que la presencia de la especie A incrementa
la tasa de crecimiento de la especie B, mientras que la presencia de la especie
B disminuye la tasa de crecimiento de la especie A. Esta sencilla definici�n
en t�rminos demogr�ficos es una fachada neocl�sica que oculta un altar churrigueresco.
Los tipos de interacci�n biol�gica que presentan las consecuencias demogr�ficas
son en extremo variados.
En este cap�tulo subdividiremos las interacciones depredador-presa en cuatro subclases: 1) depredador-presa sensu stricto, como las t�picas interacciones entre leones y gacelas, ara�as y moscas, etc.; 2) par�sito-hospedero, como las correspondientes a enfermedades virales, bacterianas, helm�nticas, etc., de los organismos en general; 3) parasitoide-hospedero, las cuales son interacciones poco conocidas entre los no especialistas, pero de la mayor importancia b�sica y aplicada y que ocurren principalmente entre insectos cuyos estadios juveniles se desarrollan dentro de alg�n hospedero (generalmente otro insecto); y, por �ltimo, 4) herb�voro-planta, como las relaciones entre las vacas y los pastos, las orugas y las plantas, etc. Aunque esta clasificaci�n puede subdividirse a�n m�s, el prop�sito del presente trabajo es proporcionar un panorama general y no una monograf�a t�cnica, por lo que las cuatro subdivisiones anteriores resultan suficientes.
Como consecuencia de las importantes diferencias biol�gicas entre los tipos
arriba mencionados, existen tambi�n diferencias metodol�gicas y te�ricas en
su estudio. A continuaci�n presentaremos brevemente la teor�a m�s general, haciendo
uso del espacio de fase, para luego particularizar en cada uno de los cuatro
tipos principales de interacci�n +- La teor�a b�sica de las interacciones depredador-presa
fue propuesta en las d�cadas de los veinte y treinta por los pioneros Alfred
Lotka y Vito Volterra. Lotka propuso sus ecuaciones haciendo una analog�a con
ciertas reacciones qu�micas, en tanto que Volterra se inspir� en un problema
sobre pesquer�as en el mar Adri�tico. Las ecuaciones, sin embargo, resultaron
id�nticas. El modelo de depredador-presa propuesto por Lotka y Volterra no tiene
m�s que una importancia hist�rica. En la actualidad, los modelos generales de
depredador-presa son modificaciones o extensiones de las ecuaciones de Lotka-Volterra.
En t�rminos generales, dichos modelos son particularizaciones del siguiente:
| Tasa de crecimiento de la presa | = | Tasa de crecimiento de la presa en ausencia del depredador | - | Tasa de mortalidad debida a la presencia del depredador |
La ecuaci�n anterior corresponde a la din�mica de la poblaci�n de las presas,
con una ecuaci�n correspondiente para los depredadores.
| Tasa de crecimiento del depredador | = | Tasa de crecimiento del depredador en ausencia de las presas de la ecuaci�n anterior | + | Incremento en la tasa de crecimiento del depredador debido a la presencia de las presas de la ecuaci�n anterior |
Los detalles biol�gicos correspondientes a un sistema o clase de sistemas en particular se traducir�n en una forma espec�fica para las ecuaciones anteriores. Por ejemplo, la ecuaci�n para las presas ser� diferente si se trata de gacelas que cuando se trata de orugas. Sin embargo, existen algunas formas generales, cualitativamente semejantes a un gran n�mero de ecuaciones particulares. Analizaremos, utilizando la herramienta del espacio de fase, una de estas formas muy generales con el objeto de derivar conclusiones cualitativas y generales.
En el espacio de fase, ubicaremos en el eje horizontal o de las X a la densidad de las presas, y por lo tanto, sus tasas de crecimiento en cualquier punto estar�n representadas por flechas en el mismo sentido horizontal. La abundancia de los depredadores se representa en el eje vertical, y sus tasas, por flechas verticales.
La isoclina cero del depredador debe tener, en general, una pendiente positiva,
puesto que resulta l�gico suponer que a mayor abundancia de las presas corresponde
una m�s alta densidad de los depredadores en el equilibrio. Sin embargo, es
muy posible que la poblaci�n de depredadores se sature de presas, es decir,
que si la densidad de presas crece por encima de cierto valor, el incremento
en la poblaci�n de depredadores llega a ser despreciable. Esto puede ocurrir
si otros factores diferentes al alimento se vuelven limitantes para los depredadores,
como, por ejemplo, el espacio para establecer territorios. Por lo tanto, la
isoclina para los depredadores tendr� una forma semejante a la de las curvas
en la gr�fica de la figura IV. 1a. Si la isoclina del depredador parte del eje
de las X, significa que se trata de un depredador espec�fico de una presa, esto
es, que no puede sobrevivir en ausencia de dicha especie de presa. En efecto,
se requiere al menos una cantidad umbral, P0 de presas (v�ase
la figura IV.1a) para que exista una cantidad positiva de depredadores. Por
el contrario, si la isoclina cero del depredador cruza el eje de las Y por encima
del origen, entonces se trata de un depredador generalista, que incluso en ausencia
de la especie de presas en cuesti�n es capaz de mantenerse en el �rea, a una
densidad D0, aliment�ndose de otras presas cuyas ecuaciones
no se consideran en el modelo (v�ase la figura IV.1b).
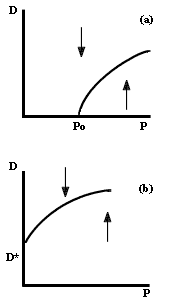
Figura IV.1. La isoclina del depredador, suponiendo un depredador especialeista
(a), y uno generalista (b).
La forma de la isoclina mencionada corresponde cualitativamente a las de un buen n�mero de modelos para el depredador. Los detalles cuantitativos var�an, pero no la forma general.
La isoclina de la presa debe tener al menos una regi�n de pendiente negativa; esto es consecuencia de que al aumentar el n�mero de presas, los factores intr�nsecos de regulaci�n de la poblaci�n de presas (mortalidad dependiente de la densidad debida a competencia, enfermedades, etc.) se acent�an, por lo que son necesarios , menos depredadores para mantener el crecimiento de las presas en cero. Por lo tanto, la parte de la isoclina de las presas m�s alejada del origen debe tener una pendiente negativa. Sin embargo, en las cercan�as del origen, la pendiente puede ser positiva o negativa, ya que existen varios fen�menos que producen una regi�n de pendiente positiva. Por ejemplo, si con bajas densidades las presas tienen problemas para encontrar pareja, un n�mero reducido de depredadores puede ser suficiente para disminuir la tasa de crecimiento de las presas hasta cero. O bien, si las presas se defienden mediante mecanismos de tipo social (todos o muchos de los individuos de un grupo participan en la defensa de �ste), entonces, cuando hay bajas densidades de presas se necesitan menos depredadores para mantener a aqu�llas en el equilibrio. Un �ltimo mecanismo que puede producir regiones de pendiente positiva es el hecho de que los depredadores pueden tener una limitada capacidad de manejo de las v�ctimas, de modo que su eficiencia sea mayor con bajas densidades de presas. Este mecanismo, denominado respuesta funcional, es de suma importancia en las interacciones depredador-presa.
En resumen, por una variedad de razones, deber�amos de esperar que la isoclina de las presas tuviera una regi�n de pendiente positiva y otra negativa; o sea una forma jorobada como aparece en la figura IV.2.
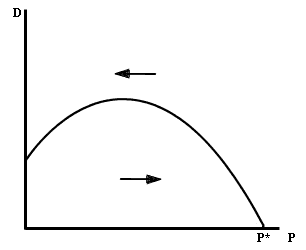
Figura IV.2. Isoclina de una presa con efecto Allee.
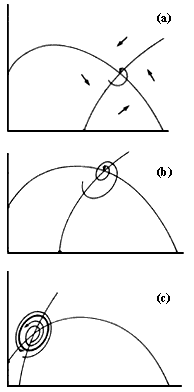
Figura IV.3. Espacios de face de una interacci�n depredador presa,
con un equilibrio no oscilatorio (a), oscilatorio (b), y un ciclo l�mite (c).
La topograf�a de flechitas que nos indica la direcci�n en la que se mover� el sistema compuesto por las presas y los depredadores aparece en la figura IV.3. Notemos que hay varias posibilidades que dependen de que el cruce de las dos isoclinas (o sea el punto de equilibrio del sistema) quede a la izquierda o a la derecha de la joroba de la isoclina de la presa. Se puede demostrar rigurosamente, para modelos particulares de este tipo, que los posibles resultados cualitativamente diferentes son tres: 1) El equilibrio es estable y las trayectorias se acercan a �l mon�tonamente, o sea, sin subidas y bajadas. Esto corresponde a la figura IV. 3a. 2) El equilibrio es estable, pero las trayectorias se acercan a �l en forma oscilatoria, es decir, a veces excediendo y a veces por debajo del valor de equilibrio (figura IV.3b). 3) El equilibrio es inestable, pero existe una regi�n en forma de curva cerrada dentro del plano que atrae a las trayectorias. Esta regi�n representa un equilibrio m�s complejo que los simples puntos que hemos encontrado hasta ahora. En otras palabras, existe dentro del plano una curva cerrada (que rodea al punto de equilibrio inestable) hacia la cual tienden las trayectorias, acerc�ndose m�s y m�s en el tiempo. Este "ciclo l�mite" (seg�n la terminolog�a matem�tica) es estable. Biol�gicamente, esta regi�n atractora corresponde a valores de la densidad de las poblaciones de depredador y su presa, que oscilan con regularidad en ciclos de baja y alta abundancia de cada uno. La estabilidad del ciclo l�mite radica en que, similarmente a lo que ocurr�a con los puntos de equilibrio, las perturbaciones en las densidades de las especies componentes del sistema tienden a desaparecer con el paso del tiempo, pues el ciclo l�mite atrae a las densidades hacia una pauta regular establecida por las ecuaciones que describen la din�mica (v�ase la figura IV.3c).
Resumiendo la teor�a expuesta, nuestro simplificado modelo matem�tico nos llevar�a a esperar que las interacciones -+ tendieran a oscilar, aunque los detalles de las oscilaciones depender�an de la forma particular de las ecuaciones. En la naturaleza se observan todas las posibilidades mencionadas.
Como ya se dijo antes, las interacciones -+ incluyen a una gran variedad de tipos biol�gicamente muy distintos. Posiblemente las m�s conocidas, o m�s obvias, son las depredador-presa en un sentido estricto. �Qui�n no ha visto a un gato acechando a un p�jaro? �O a una mosca o abeja presa en la tela de una ara�a? �O incluso, en esta �poca de televisi�n y cine, magn�ficas y sorprendentes im�genes de leones o cheetas en el momento mismo de capturar a su presa? Como dijera el c�lebre ec�logo Charles Elton, posiblemente la forma de muerte m�s com�n para los animales en la naturaleza sea la de ser devorados por alg�n otro. Los depredados en general son animales (aunque existen tambi�n plantas) de un tama�o similar al de sus presas, y el acto de la depredaci�n, cuando es exitoso, culmina invariablemente con la muerte de la presa.
Claramente, los depredadores deben de representar un papel importante en la econom�a de la naturaleza. Hoy d�a, las interacciones -+ se consideran sumamente importantes para la estructura de las comunidades, por el hecho de que los depredadores pueden tanto impedir la presencia de algunas especies, como facilitar la de otras, tal como ilustra apropiadamente el trabajo realizado por Richard Paine. En la d�cada de los sesenta, Paine realiz� un experimento, considerado ahora cl�sico, que consisti� en eliminar al depredador m�s importante (una estrella marina del g�nero Pisaster) de una comunidad de invertebrados marinos de la zona de intermarea. Inesperadamente, el resultado fue que el n�mero de especies presente disminuy� de manera brusca de cerca de quince a ocho. La raz�n de este decaimiento es que la estrella de mar manten�a las poblaciones de muchos potenciales competidores muy por debajo de los niveles en los que la competencia se habr�a hecho significativa. Al eliminarse al depredador, algunas especies de presas ejercieron una interacci�n competitiva tan severa en las otras que las excluyeron por completo de la comunidad. Sin embargo, el efecto de la depredaci�n no siempre es igual. Un depredador que no distinga entre sus presas puede producir un efecto reductor de la diversidad de la comunidad, al disminuir por parejo las densidades de todas o muchas de las especies.
Hay muchas formas de ser depredador, as� como modos de defenderse. Desde tiempo antiguo, el hombre se ha admirado de las maneras que los animales, reales o m�ticos, tienen de atacar o defenderse. Como ejemplo, tomemos este pasaje de la Historia natural de Cayo Plinio Segundo, tal como lo presenta don Francisco Hern�ndez:
| Mira pues el drag�n por d�nde va el elephante a sus pastos, y arr�jase desde un �rbol alto a �l. Entiende el elephante que no podr� valerse contra sus ataduras y ans� busca �rboles y pe�ascos, donde estreg�ndose lo mate. Gu�rdanse los dragones desto y por lo tanto les atajan los pasos con la cola. Desatan los elephantes con la trompa su �udo, mas ellos les meten la cabeza en las narices e impidi�ndoles el haliento les roen aquellas partes tern�simas. |
No necesitamos buscar ejemplos mitol�gicos para descubrir la variedad de formas de la interacci�n depredador-presa. Un gran n�mero de especies de depredadores son del estilo denominado emboscadores, o sea organismos que acechan inm�viles a sus presas hasta tenerlas muy cerca, o bien hasta que caen en alguna trampa. Ejemplos de depredadores que cazan con trampa son las ara�as que tejen redes, las hormigas le�n que construyen embudos en la arena, las plantas carn�voras que producen sustancias pegajosas, el pejesapo que simula un gusano enfrente de la boca para atraer pececillos hambrientos, etc. Acechadores, que aguardan hasta que la presa est� cerca, son algunos felinos (jaguares, gatos, leopardos), algunas serpientes, ara�as como los thom�sidos, muchos reptiles (camaleones), etc. Otra amplia categor�a de depredadores son los que buscan de manera activa a sus presas, ya sea colectivamente, como los lobos y a veces los leones, o solitariamente, como muchas aves de presa, peces y mam�feros carn�voros, ara�as como los salt�cidos, etc�tera.
Por su parte, las presas tambi�n han desarrollado algunos estilos t�picos de defensa, como son el ocultarse (insectos cr�pticos), los movimientos �giles y r�pidos (gacelas, aves), la posesi�n de armaduras (pangol�n, puerco espines, algunos moluscos) y defensas qu�micas internas (toxicidad o mal sabor) o externas (tinta de pulpos y calamares, mal olor en zorrillos, etc�tera).
A lo largo de la historia, la selecci�n natural favorece a aquellos depredadores que son m�s eficientes y al mismo tiempo a las presas que evitan mejor a sus enemigos. Hay muchos casos en los que a cada nueva adaptaci�n del depredador sigue (en el tiempo evolutivo) una contraadaptaci�n de las presas, a la cual subsecuentemente responden los depredadores. A este fen�meno se le llama coevoluci�n, y es claro que el proceso por el cual depredadores y presas desarrollan cada vez mejores adaptaciones como una respuesta a las de su contraparte no tiene por qu� acabar. Esta "carrera armamentista", como ha sido llamada por Dawkins, debe de producir cada vez m�s finas adaptaciones, y en efecto, un cat�logo de adaptaciones para resolver tanto el problema de depredar como el de no ser depredado ser�a interminable. Veremos ahora unos pocos ejemplos ilustrativos.
Una interacci�n t�pica entre un gremio de depredadores y otro de presas es aquella que se puede observar entre las aves insect�voras y sus insectos presas, en particular mariposas. Una de las v�as evolutivas que las mariposas han seguido y que las hacen presas menos f�ciles es la cripsis. La cripsis es el fen�meno por el cual una especie se asemeja a alguna parte de su h�bitat (suelo, piedras, hojas, varas, etc�tera). Existen mariposas y palomillas asombrosamente semejantes a su entorno. Muchas especies del g�nero tropical Anea imitan no s�lo la coloraci�n de las hojas secas, sino tambi�n las venaciones y marcas de dichas hojas, incluyendo las manchitas producidas por los hongos. Una Anea parada sobre el umbr�o suelo de una selva resulta casi imposible de diferenciar de las hojas semiputrefactas.
Por supuesto, la coloraci�n cr�ptica no est� limitada a las mariposas. Los famosos insectos palo no s�lo poseen una coloraci�n, inclusive tienen una forma que los asemeja notablemente a su entorno. Los camaleones africanos, los pulpos y muchos peces, son capaces de cambiar de color por medios fisiol�gicos (dilataci�n y contracci�n de peque�as estructuras coloreadas), con el resultado de que en minutos pueden confundirse con el medio.
Otros organismos se "disfrazan" activamente, para asemejarse a su medio ambiente. Tal es el caso de los cangrejos marinos de la familia Majidae. Muchas especies dentro de esta familia cubren su caparaz�n con algas, peque�as esponjas, an�monas, piedrecillas, etc. El proceso es complicado y requiere de la obtenci�n del "decorado" y su preparaci�n, para lo cual el cangrejo mastica un extremo del alga, esponja o lo que sea, hasta dejarlo rugoso. Posteriormente, se coloca la pieza del decorado sobre el caparaz�n, y para ello el cangrejo cuenta con unos peque�os ganchitos, a manera de vellosidad, que sirven para sujetar la decoraci�n. Un cangrejo bien disfrazado reposando sobre un sustrato rocoso es pr�cticamente invisible para el ojo humano, y probablemente tambi�n para los depredadores tales como pulpos, una variedad de peces, nutrias marinas, etc. Pero el decorado tambi�n sirve para que las presas potenciales se acerquen al cangrejo, por lo que su funci�n es doble: protege al organismo de sus depredadores y lo oculta de sus presas.
La cripsis es una adaptaci�n obvia en contra de los depredadores, pero hay algunas m�s sutiles. Existe otra v�a evolutiva que da respuesta al problema de los depredadores en una forma opuesta a la cripsis. La llamada coloraci�n aposem�tica consiste en dibujos y colores (y pautas de conducta) muy conspicuos que avisan al depredador que la supuesta presa no es tal. Las larvas de muchos insectos se alimentan de plantas que contienen sustancias altamente t�xicas para las aves o los mam�feros. Muchas especies de insectos son capaces no s�lo de tolerar dichas toxinas, sino de acumularlas durante su desarrollo hasta el estado adulto, resultando de ello que el insecto es t�xico o de sabor repugnante para un depredador potencial. Experimentalmente es posible demostrar que un p�jaro que prueba una de estas mariposas "protegidas" aprende a reconocer el dise�o de coloraci�n de la presa protegida y rechaza subsecuentemente ofrecimientos de mariposas de la misma especie. Los trabajos de Lincoln y Jane Brower indican que un ave puede recordar el dibujo de la mariposa t�xica hasta por unos tres meses despu�s de su primera experiencia.
Obviamente, un dise�o de coloraci�n conspicuo y f�cil de recordar (desde un punto de vista de ave) debe proteger mejor a las especies t�xicas; y en efecto, en una selva alta tropical, muchas de las especies de mariposas protegidas (por ejemplo de las familias Heliconidae e Ithomidae) presentan pautas de coloraci�n brillantes y muy visibles, asociadas a un vuelo lento y descuidado.
Posiblemente a estas alturas m�s de un lector ya se haya planteado la pregunta de c�mo puede evolucionar la coloraci�n aposem�tica, puesto que los beneficiados por ella son siempre los individuos no atacados. Ense�arle a un depredador que una cierta pauta de coloraci�n corresponde a cosas-que-no-son-comida, muy posiblemente le cueste la vida al primer insecto vistoso, y por lo tanto los genes responsables de la coloraci�n vistosa no deber�an de propagarse en la poblaci�n. La soluci�n te�rica a esta aparente paradoja yace en el concepto de "selecci�n de parentesco, propuesto originalmente por Maynard-Smith hace unas tres d�cadas. El punto est� en percibir que cualquier gene de un individuo tiene ciertas probabilidades de encontrarse en otros individuos. Dichas probabilidades son m�s altas mientras m�s cercano es el parentesco entre los individuos en cuesti�n. Por lo tanto, una mariposa muy vistosa que es devorada por un ave, aunque ya no generar� directamente copias de sus genes, puede muy bien proteger a aquellas copias residentes en sus parientes y que resultar�n promovidas por el hecho de que el ave evitar� en lo sucesivo la pauta de coloraci�n asociada con el sabor desagradable. Es obvio que para que este mecanismo trabaje se requiere que los individuos de una localidad tengan un grado alto de consanguineidad. Tenemos entonces una explicaci�n plausible (aunque a�n hipot�tica) de la existencia de la coloraci�n vistosa en mariposas t�xicas.
Una tercera adaptaci�n, a�n m�s sutil que las mencionadas antes, para evitar a los depredadores, es la que presentan muchas mariposas vistosas pero que no son t�xicas ni de sabor desagradable. Existen especies que son perfectamente aceptables como alimento para sus depredadores, y que adem�s se parecen en color e incluso en pautas de vuelo a especies aposem�ticas protegidas por su toxicidad. El resultado es que las mariposas no t�xicas son evitadas por los depredadores que han aprendido de las t�xicas a rechazar la pauta de coloraci�n com�n a ambas. Este fen�meno, denominado mimetismo de Bates, se encuentra particularmente bien representado en los tr�picos, donde el parecido entre algunas especies llega a ser casi perfecto.
Por supuesto, los depredadores pueden incrementar su tasa de captura de presas siguiendo v�as evolutivas del tipo de las descritas para presas. Por ejemplo, la piel de leones, tigres y leopardos se asemeja bastante a la coloraci�n del h�bitat preferido por estas especies. Existen otros casos, menos conocidos, de depredadores espectacularmente bien ocultos a la vista de sus presas, como muchas ara�as y campamochas cr�pticas. Tambi�n se conocen ejemplos de depredadores que atraen a sus presas, como la campamocha africana Hymenopus coronatus, la cual tiene color y forma de flor y atrapa a los insectos que, enga�ados, acuden a buscar n�ctar o polen. Esta clase de mimetismo (llamado de Peckham) ha sido adoptado por grupos completos de ara�as (los Misum�nidos) y de chinches (los Phym�tidos).
Muchos estilos de depredaci�n implican haber resuelto el problema de detectar a presas peque�as, muy m�viles o simplemente activas durante la noche, cuando la luz disponible es pobre o inexistente. Los ojos de muchos depredadores est�n finamente adaptados para resolver este problema: por ejemplo, la retina de los felinos que acechan de noche puede abrirse para ocupar casi toda el �rea disponible del ojo y dejar pasar un m�ximo de luz. Las v�boras de cascabel cuentan con la capacidad de "ver" el calor, y por lo tanto pueden detectar a sus presas en la oscuridad por contraste con las �reas fr�as inertes. Los �rganos termosensibles de las v�boras de cascabel est�n localizados en peque�as cavidades abajo y un poco adelante de los ojos, y son tan sensibles que empiezan a responder ante cambios de temperatura �desde tres mil�simas de grado cent�grado!
El sentido del o�do tambi�n se ha desarrollado mucho en algunos depredadores. Por ejemplo, los b�hos o tecolotes localizan a sus presas (ratones, principalmente) por el ruido que �stas hacen al desplazarse por el suelo, muchas veces en una oscuridad total. El b�ho debe determinar el �ngulo y la posici�n horizontal a la que se encuentra la presa. Como Eric Knudsen ha demostrado, la anatom�a de la cara de los b�hos (en particular los de la familia Tytonidae) est� exquisitamente adaptada para los fines de percibir e interpretar los tenues sonidos producidos por los movimientos de un rat�n en la hojarasca. La cara redonda y cubiertade plumas muy densas y apretadas es un excelente reflector de sonidos de alta frecuencia. La estructura �sea del cr�neo de los b�hos es tal que el o�do derecho es m�s sensible a los sonidos de alta frecuencia que llegan de arriba, y el izquierdo a los sonidos que llegan de abajo. La diferencia de intensidades provee al b�ho de informaci�n sobre la elevaci�n de una fuente de sonido y sobre su posici�n horizontal.
Tanto los b�hos como los cr�talos est�n delicadamente adaptados para encontrar a sus presas. Sin embargo, posiblemente el conjunto de adaptaciones m�s fascinantes para resolver el problema de detectar presas peque�as en la oscuridad es el ejemplificado por el "sonar" de los murci�lagos y analizado por Richard Dawkins para ilustrar los fant�sticos resultados de los procesos selectivos.
En los a�os cuarenta se descubri� que muchas especies de murci�lagos eran capaces de detectar la presencia de objetos y presas aun en completa oscuridad. Desde esa fecha se ha avanzado mucho en el entendimiento de los mecanismos de "ecolocalizaci�n". La idea b�sica es muy simple: el murci�lago emite sonidos de muy alta frecuencia (y por lo tanto en su mayor�a inaudibles para nosotros) que al reflejarse en los obst�culos enfrente de �l (por ejemplo, los insectos que constituyen su alimento), son detectados por el animal y le informan de la posici�n, el tama�o e incluso la velocidad y direcci�n del movimiento de la presa. �sta es la misma idea del radar y del sonar, con la salvedad de que la selecci�n natural, actuando sobre los murci�lagos, la desarroll� posiblemente unos 50 millones de a�os antes que las invenciones humanas. Para que un sistema de ecolocalizaci�n a�rea funcione bien, es necesario resolver un gran n�mero de problemas t�cnicos. Mencionaremos solamente dos de ellos: el primero radica en que el sonido, al ser emitido por su fuente, pierde intensidad de manera muy r�pida (cuadr�ticarnente), es decir que si la onda sonora avanza dos metros, la intensidad del sonido se reduce no a la mitad, sino a la cuarta parte; si son cinco metros, la intensidad se reduce a la vig�sima quinta parte. Como, adem�s, el sonido emitido por el murci�lago debe de reflejarse (tal vez desde un blanco de pocos mil�metros cuadrados de superficie) y recorrer una distancia similar a la del viaje de ida, tenemos que el sonido que regresa al animal puede ser de una intensidad baj�sima. Por lo tanto, un murci�lago con un ecolocalizador eficiente debe: 1) emitir sonidos de muy alta intensidad, y 2) poseer receptores de una grand�sima sensibilidad. Tenemos entonces dos requerimientos que podr�an parecer contradictorios, porque los gritos (ultras�nicos) que emite el murci�lago cada fracci�n de segundo deben de ser ensordecedores, literalmente, para sus hipersensibles receptores. Este mismo problema se present� a quienes dise�aron el radar, y fue resuelto mediante un sistema que desconecta el receptor un instante antes de que se emita el pulso de radio. Pues bien, el mismo sistema se encuentra en muchas especies de murci�lagos, las cuales tienen ciertos m�sculos adosados a los huesecillos auditivos que trasmiten el sonido desde el t�mpano. Un instante antes de que el murci�lago grite, estos m�sculos se contraen y detienen el paso del sonido, evitando as� el ensordecimiento. Hay especies que realizan este proceso de "prendido y apagado" hasta cincuenta veces por segundo.
Otro importante problema adaptativo es conocer la velocidad a la que se mueve la presa. Para resolverlo, muchas especies de murci�lagos tienen mecanismos que se basan en el llamado efecto Doppler. Todos conocemos, aun sin saber su nombre, este efecto. Cuando una ambulancia o patrulla viene hacia nosotros, la sirena se escucha con un tono agudo, pero cuando la ambulancia pasa y empieza a alejarse, el tono cambia y se hace m�s grave. La raz�n es que las sucesivas compresiones en el aire que constituyen la onda sonora se reciben m�s frecuentemente cuando viajan hacia nosotros que cuando se alejan, y la frecuencia es lo que da la agudez del sonido. Cuando el murci�lago recibe un eco de un objeto que se aproxima a �l, cada percepci�n del eco es m�s aguda que la anterior. Cuando, por el contrario, el objeto se aleja del murci�lago, el eco de cada emisi�n de sonido es de tono m�s bajo. Comparando las diferencias de tono, el cerebro y el aparato auditivo de las especies de murci�lagos que cuentan con este mecanismo pueden calcular la velocidad relativa de los objetos que se encuentran enfrente de ellos.
Existen otras maravillosas adaptaciones en el mecanismo de ecolocalizaci�n de los murci�lagos, pero para terminar la secci�n s�lo diremos que muchas presas de los murci�lagos han respondido evolutivamente con sus propias contradefensas, desde un o�do capaz de percibir el sonar del murci�lago para seguir la conducta de evasi�n adecuada (como en algunas mariposas nocturnas), hasta la emisi�n de sonidos que interfieren con el ecolocalizador del atacante (algunas ranas tropicales). La carrera coevolutiva entre depredadores y presas nunca termina, mientras las especies competidoras permanezcan en el juego.
El siguiente tipo de interacci�n -+ del que hablaremos, es el de los parasitoides y sus hospederos. Los parasitoides son insectos, en su mayor�a avispas (en su sentido taxon�mico, esto es, himen�pteros) y moscas. Los parasitoides depositan sus huevecillos, o a veces a sus larvas, dentro o muy cercanamente a sus hospederos, quienes habitualmente son otros insectos y a veces ara�as. El parasitoide crece, se desarrolla dentro del hospedero, y acaba mat�ndolo. De un insecto parasitado puede surgir de uno a muchas decenas de parasitoides, dependiendo de la especie de �ste, el tama�o del hospedero, etc�tera.
A diferencia de lo que ocurre con los depredadores "verdaderos", los parasitoides son totalmente desconocidos para el com�n de la gente, al grado de que se pudiera pensar que son meras curiosidades de la biolog�a sin mayor importancia ecol�gica o econ�mica. Nada podr�a estar m�s alejado de la verdad. Los parasitoides, num�ricamente, son uno de los grupos ecol�gicos m�s importantes, compuesto por no menos de unas ciento cuarenta mil especies. Es muy probable que la mayor�a de los insectos herb�voros (importantes plagas agr�colas) se mantengan en niveles poblacionales bajos como consecuencia de la acci�n de los parasitoides. La gran cantidad de ejemplos exitosos de control biol�gico de plagas utilizando parasitoides (algunos de ellos se presentar�n en el cap�tulo final) subrayan la importancia que esta interacci�n tiene para determinar los n�meros de muchas especies de insectos. En efecto, no es raro que la introducci�n de un parasitoide reduzca en varios �rdenes de magnitud la densidad de la presa, con el resultado de que ambas pueden aparecer como especies raras y poco importantes, y solamente el conocimiento de la historia de la interacci�n podr�a demostrar lo contrario.
El "frijolarium", ya sea f�sico o programado que se describe en el ap�ndice, puede ilustrar la tendencia oscilatoria de las interacciones -+, pero no constituye un buen modelo del fen�meno descrito porque no se puede representar bien la heterogeneidad medioambiental, que resulta esencial para entender las relaciones din�micas entre parasitoides y hospederos.
Para ilustrar la ca�da de una poblaci�n de hospederos como consecuencia de la introducci�n de parasitoides se puede recurrir a un modelo simple, que considera los ataques de los parasitoides sobre sus hospederos como una variable aleatoria con alg�n tipo de distribuci�n "agregada". Esto es, como consecuencia de la heterogeneidad medioambiental, algunos hospederos tienen mucha m�s alta probabilidad de ser atacados que otros. Si representamos la frecuencia de hospederos con x ataques como f(x), entonces, si en un tiempo t hab�a n(t) hospederos, se librar�n simplemente n(t) f(�). Si cada uno de ellos produce en promedio F descendientes, tenemos que en el tiempo t + 1 habr�:
|
|
|
Similarmente, si f(�) hospederos no fueron atacados, entonces el reciproco 1 - f(�) s� lo fue, y como para muchas especies de parasitoides ocurre que no se dan ataques m�ltiples, o bien, el n�mero que emerge de un hospedero no depende del n�mero de ataques, sino de factores como el tama�o del hospedero, etc., tenemos que cada hospedero atacado producir� en promedio una cantidad a de nuevos parasitoides. Denotando los parasitoides en el tiempo t con p(t), tenemos entonces:
|
|
|
Ahora escogeremos a la distribuci�n llamada "binomial negativa" para representar el n�mero de ataques. Esto es, supondremos que la probabilidad de encontrar hospederos con cero ataques, con un ataque, con dos ataques, etc., est� dada por la f�rmula de la binomial negativa. Esta suposici�n se hace por razones emp�ricas: se sabe que la distribuci�n de ataques en muchos casos es agregada, como la distribuci�n binomial negativa. Sustituyendo, obtenemos:
|
|
|
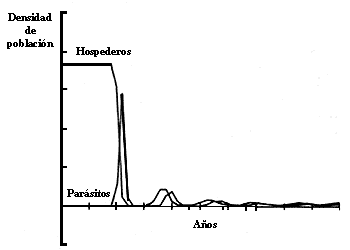
Figura IV.4. Din�mica temporal de una interacci�n parasitoide-hospedero
con una distribuci�n de ataques contagiosa. En este modelo, la introducci�n
del parasitoide produce un abatimiento de la poblaci�n hospedera a un 2% del
valor original.
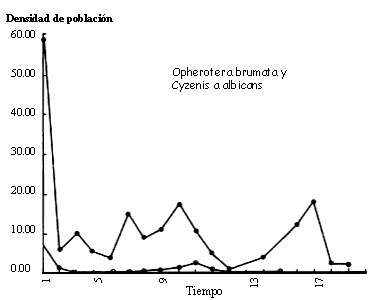
Figura IV.5. Valores observados para la interacci�n entre Operophtera
brumata y Cyzenis Albicans. Datos de Varley, G., G. Gradwell, y M.
Hasell (1973). Blackwell Press.
donde k es el �ndice de agregaci�n (k < 1 implica agregaci�n en la distribuci�n. k>>1 corresponde a una distribuci�n de Poisson, o sea al azar) y b es un �ndice de la capacidad del parasitoide para encontrar a los hospederos. Simulando este modelo en una computadora, se obtienen gr�ficas como las de la figura IV.4, en las que se puede apreciar el abatimiento en el n�mero de hospederos que pueden darse por la acci�n de parasitoides.
Otra consecuencia importante del uso del modelo binomial negativo es que con �l resulta f�cil imitar una caracter�stica importante de las interacciones parasitoide-hospedero observadas en el campo, a saber: que las densidades de equilibrio son muy bajas y rara vez oscilan (v�ase la figura IV.5). Si se supone que el medio ambiente es homog�neo, lo cual implicar�a una distribuci�n de ataques de tipo Poisson, la k del modelo resulta grande y los equilibrios tienden a ser inestables y darse s�lo con alta densidad. Si se modela la heterogeneidad ambiental por medio de la disminuci�n de la k para simular una distribuci�n agregada de ataques, se obtienen equilibrios mucho m�s parecidos a los que ocurren en la naturaleza.
Las adaptaciones producidas por esta interacci�n no pod�an dejar de ser m�ltiples y fascinantes. Al igual que para los depredadores, el problema de localizar a los hospederos es central para los parasitoides. Muchos de ellos localizan primero al microh�bitat o la planta de alimentaci�n de sus hospederos, para luego buscarlos. Otros son atra�dos por los olores que la planta produce al ser devorada por el hospedero, y hay otros capaces de detectar el ruido que hace el hospedero al masticar las hojas. Ciertas avispas parasitoides de la familia de los Ichneum�nidos atacan a hospederos que se ocultan bajo la corteza de los �rboles. La hembra localiza primero el sitio de la corteza bajo el cual se encuentra su v�ctima, y entonces introduce el ovipositor atravesando a veces m�s de un cent�metro de corteza dura, hasta dejar sus huevecillos en el cuerpo del insecto.
Por supuesto, tambi�n en los hospederos se han generado adaptaciones como una respuesta a las presiones selectivas producidas por los parasitoides. Existen soluciones morfol�gicas, tales como largas sedas y pelos que cubren a las larvas de muchas mariposas y palomillas, y que dificultan a los parasitoides efectuar la oviposici�n. Muchos insectos del orden de los Hom�pteros se cubren de sustancias c�reas o espumas que tambi�n pueden servir de protecci�n contra parasitoides. En otros casos, la respuesta es de tipo conductual. Por ejemplo, hay larvas de mariposas que al percibir la cercan�a del parasitoide se dejan caer de la hoja, manteniendo un "cable de seguridad" de seda adherido a la misma. Pasados unos cuantos minutos, la larva regresa a su hoja trepando por el cable. Finalmente. existen defensas inmunol�gicas, que permiten al hospedero combatir a su enemigo cuando �ste ya se halla dentro del cuerpo del primero.
Michael Rosenzweig, analizando un modelo del tipo de los de la figura IV.3, propuso que la coevoluci�n entre los depredadores y sus presas pod�a estabilizar el r�gimen oscilatorio como consecuencia de un incremento en la capacidad defensiva de las presas. Esta hip�tesis se ve claramente apoyada por los experimentos cl�sicos de Francisco Pimentel, quien encontr� que, en un sistema de laboratorio, las violentas oscilaciones de moscas caseras y sus parasitoides (Nasonia vitripennis) se amortiguaban hasta desaparecer al ser seleccionados genotipos de moscas resistentes al ataque de los parasitoides.
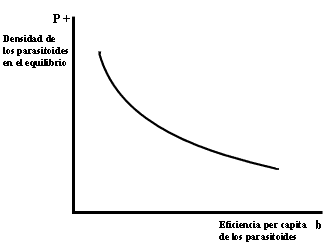
Figura IV.6. Disminisi�n de la poblaci�n de parasitoides como consecuencia
de aumentos en eficiencia per capita.
Decir que los hospederos son m�s resistentes equivale a se�alar que los parasitoides son menos efectivos como tales. Ahora bien, ser�a de esperarse que la selecci�n natural favoreciera depredadores cada vez m�s eficientes, puesto que �stos son los que con mayor probabilidad deber�an pasar sus genes a su progenie. Es decir, se esperar�a que la selecci�n natural actuara para incrementar el par�metro b de la ecuaci�n 4.3. Si se simula esto en el modelo, se obtiene el resultado de que aumentos en la eficiencia de los parasitoides redundan en una disminuci�n conjunta de las poblaciones de hospederos y parasitoides, con el consecuente incremento del riesgo de extinguirse (v�ase la figura IV.6). Por lo tanto, aunque en teor�a esperar�amos que la selecci�n natural en un corto plazo favoreciera la evoluci�n de depredadores m�s eficientes, en poblaciones de depredadores muy cerradas, sin flujos migratorios, el grupo completo podr�a extinguirse como consecuencia de la selecci�n individual. La hip�tesis del "depredador prudente" entonces sugiere que la selecci�n deber�a de favorecer depredadores con eficiencias intermedias. Esta hip�tesis depende sobre todo de la suposici�n de que la poblaci�n de depredadores es cerrada a la emigraci�n, cosa que posiblemente no sea com�n entre los parasitoides. En la siguiente secci�n veremos algunos candidatos m�s veros�miles para el t�tulo de depredadores prudentes.
Las interacciones -+ de las que hablaremos ahora son las de los par�sitos y sus hospederos. Los par�sitos son organismos peque�os en relaci�n con sus hospederos, que no necesariamente mueren a consecuencia de la interacci�n; normalmente, provienen de l�neas evolutivas muy diferentes a las de sus hospederos, y viven en una relaci�n muy estrecha (simbi�tica) con el hospedero del cual obtienen su alimentaci�n.
Gran cantidad de estas interacciones han sido muy bien estudiadas, en particular aquellas que afectan al ser humano (diarreas, infecciones respiratorias, peste bub�nica, SIDA, paludismo, etc.), aunque s�lo recientemente los ec�logos se han dedicado a estudiar las enfermedades parasitarias como un factor importante en la din�mica de las poblaciones naturales.
El modo de vida paras�tico es uno de los m�s numerosos que se conocen, y resulta muy dif�cil hacer generalizaciones que sean v�lidas para virus, bacterias, protozoarios, nem�todos, helmintos, �caros, hongos, etc. Por lo tanto, hay que subdividir y tratar de definir subclases m�s o menos homog�neas dentro del universo de las interacciones par�sito-hospedero. Una subdivisi�n que ha resultado �til para los ec�logos de poblaciones es la presentada por Roy Anderson. Se separa a los par�sitos en dos grandes grupos: en el primero se encuentran especies por lo general microsc�picas (virus, bacterias) y que inducen en sus hospederos una reacci�n inmunol�gica m�s o menos permanente. En los modelos matem�ticos correspondientes se divide a la poblaci�n de hospederos en clases: susceptibles, inmunes y enfermos; cada clase tiene su propia din�mica, por lo qu� el espacio de fase de estos modelos rara vez se puede representar en el plano, como suced�a en casos anteriores. En el segundo grupo se incluyen par�sitos de tama�o macrosc�pico (nem�todos, helmintos, �caros, etc.) que no generan reacciones inmunes notables, ni a largo plazo. Sus modelos matem�ticos a menudo incluyen t�rminos para representar la distribuci�n estad�stica de los par�sitos dentro de sus hospederos.
Obviamente, la anterior es una clasificaci�n arbitraria y llena de excepciones, pero preserva un cierto grado de realismo y resulta muy conveniente en la medida en que separa tipos extremos de parasitismos con sus correspondientes tipos de modelos. Por otra parte, hay una serie de propiedades ecol�gicas comunes a ambos tipos de enfermedades y que se hacen evidentes como consecuencia de la modelaci�n matem�tica. Un concepto importante es la llamada tasa reproductiva b�sica de la enfermedad, denotada por R. Para el primer grupo de enfermedades (como las virales y las bacrianas), R representa el n�mero promedio de infecciones que un hospedero infectado genera durante el tiempo de su infecci�n. Claramente, si la infecci�n ha de prosperar en la poblaci�n, cada enfermo debe, en promedio, de infectar a cuando menos otro susceptible. Esto es, R > 1. Por ejemplo, Roy Anderson ha calculado que la R para el paludismo en cierta localidad de �frica Oriental es de 39, es decir que un enfermo infecta en promedio a otros 39 antes de sanar. La R para la rubeola en Cirencester, Gran Breta�a, era de alrededor de 14. Para el grupo de enfermedades producidas por nematodos, platelmintos, etc., R se interpreta como el n�mero de par�sitos que alcanzan la edad reproductiva, producidos por un par�sito promedio. Nuevamente, para que la infecci�n prospere se requiere que R sea mayor que uno. Como veremos con un poco m�s de detalle en el �ltimo cap�tulo, el estudio de los factores que afectan a R es de gran utilidad para el dise�o de estrategias de control de las enfermedades.
�Cu�les pueden ser los resultados de las mutuas presiones selectivas entre par�sitos y hospederos? Se han escrito monograf�as dedicadas exclusivamente a explorar y responder esta pregunta. Dado el car�cter de este libro, tendremos que limitarnos a presentar unos cuantos ejemplos y mencionar algunas generalizaciones. Principiaremos retomando la pregunta de la secci�n anterior. �Podemos esperar la evoluci�n de "par�sitos prudentes"? En este caso nos referir�amos a organismos poco virulentos, capaces de coexistir con sus hospederos sin matarlos. Se mencion� antes que uno de los requisitos importantes para esperar la evoluci�n de los depredadores prudentes era una gran cohesi�n en el grupo, de tal forma que los grupos formados por depredadores muy efectivos tuvieran una alta probailidad de desaparecer al acabar con sus presas antes de poder difundir los genes responsables de la efectividad.
En el caso de los par�sitos, la selecci�n de grupos se facilita. Una de las propiedades ecol�gicas esenciales de la relaci�n par�sito-hospedero es su estructura en mosaico. En otras palabras, los par�sitos viven en peque�as islas de medio ambiente favorable (sus hospederos), rodeadas de un mar de medio inh�spito (el exterior del hospedero). Si la isla desaparece (el hospedero muere) sin que el grupo de par�sitos contenidos en �l haya invadido otras islas (infectado otros hospederos), cualquier novedad evolutiva producida por el grupo desaparecer�. Por lo tanto, entre los par�sitos es donde deber�amos encontrar los mejores ejemplos de depredadores prudentes, que en este caso aparecer�an como par�sitos poco virulentos. Existen algunos casos que apoyan esta idea. Por ejemplo, se han realizado experimentos en los que se manten�an creciendo poblaciones de Escherichia coli junto con algunos de sus virus par�sitos (los fagos T3, T4 y T7). Un resultado general de estos experimentos era que, pasado un tiempo de iniciada la interacci�n, la bacteria se hac�a m�s resistente y las l�neas de bacteri�fagos menos virulentas. Por otro lado, un buen ejemplo de campo de este fen�meno lo encontramos en la myxomatosis del conejo. La invasi�n de conejos que sufrieron los australianos a principios de siglo fue controlada introduciendo un virus, el myxoma, el cual result� totalmente eficaz para abatir la plaga de conejos. Estudios subsecuentes indican que no s�lo los conejos que sobrevivieron son m�s resistentes que sus progenitores a la cepa original de myxoma, sino que las cepas del virus actualmente prevalecientes en el campo son menos virulentas.
Por supuesto, el hacerse menos virulentos no es la �nica adaptaci�n de los par�sitos frente al problema de tener que explotar islas de recursos muy concentrados e inestables temporalmente. Otra adaptaci�n consiste en presentar formas de resistencia que permitan al organismo mantenerse vivo e inactivo mientras resida en el mar de medio inh�spito que rodea a sus hospederos. Por ejemplo, hay nem�todos que pueden permanecer inactivos, fuera de sus hospederos, hasta por unos 30 a�os; las cercarias de algunos trem�todos pueden sobrevivir hasta cinco a�os; las esporas de un hongo par�sito de las cigarras peri�dicas, que aparecen de manera sincronizada cada 17 a�os exactamente, deben permanecer vivas por ese periodo de tiempo.
El lector interesado en el ampl�simo tema de la parasitolog�a deber� remitirse a las obras especializadas para profundizar en este campo fascinante.
Como �ltimo subgrupo de interacciones dentro de la categor�a -+, veremos las relaciones entre los herb�voros y las plantas. Muchas de estas relaciones son perfectamente aparentes (no se necesita ser agricultor para apreciar la importancia de herb�voros como las vacas y los borregos, o de las plagas de los cultivos de los cuales nos alimentamos). Estrictamente hablando, los herb�voros deber�an de subdividirse por lo menos en tantas categor�as como los depredadores. En realidad, casi lo �nico en com�n entre vacas, pulgones y chapulines (para no mencionar nem�todos vegetarianos, agallas, virus, hongos y otros grupos a�n menos conocidos), es que obtienen su alimento de las plantas. Sin embargo, nuestros esfuerzos por clasificar, catalogar y separar deben de tener un l�mite, por lo que sin hacer mayores especificaciones en esta secci�n nos referiremos principalmente a insectos y vertebrados que consuman materia vegetal sobre plantas vivas. Aun esta restricci�n incluye un universo m�s que satisfactoriamente rico. Alrededor de unas trescientas setenta mil especies descritas corresponden a estos grupos, y m�s del 98% de todos estos herb�voros son insectos (principalmente mariposas y palomillas, escarabajos, chinches, moscas y chapulines). Debe insistirse en especies descritas porque es seguro que a�n quedan varios millones de especies por describir (esto es, por registrarse oficialmente ante la ciencia) dentro de grupos como los insectos, los �caros y los nem�todos.
Como vimos antes, tanto los depredadores como los parasitoides matan a sus presas. En cambio, los par�sitos pueden matar, da�ar m�s o menos severamente, o incluso no afectar los par�metros demogr�ficos de sus hospederos. Algo semejante ocurre con los herb�voros: el da�o sufrido por una planta como consecuencia de la acci�n de los herb�voros puede ser desde pr�cticamente nulo hasta la defoliaci�n total y la muerte; en consecuencia, para su estudio te�rico, los sistemas herb�voro-planta se han subdividido en interactivos y no interactivos. Los �ltimos son aquellos en los que los herb�voros no afectan la tasa de renovaci�n de la vegetaci�n, mientras que en los interactivos s� se afecta dicha tasa. Es posible que la mayor�a de los insectos herb�voros pertenezca a la categor�a de los no interactivos. Esta afirmaci�n se basa en extrapolaciones de unos pocos casos muy bien estudiados (el helecho Pteridium aquilinum, el pasto Holcus mollis y las plantas llamadas aves del para�so, Heliconia Spp) en los cuales la gran mayor�a de los herb�voros encontrados sobre las plantas son muy escasos, y casi todo el da�o que reciben se debe a una o muy pocas especies de insectos abundantes.
Los principales factores que mantienen bajas las poblaciones de herb�voros son tres: en primer lugar, la acci�n de los enemigos naturales, principalmente parasitoides. El caso de los pl�tanos (Musa Sp.) en los tr�picos americanos ilustra bien este punto. En condiciones naturales, algunas especies de mariposas son los principales herb�voros y el da�o muy rara vez resulta considerable. Sin embargo, en las grandes plantaciones comerciales que a menudo son rociadas con insecticidas para disminuir a�n m�s los da�os, suele ocurrir que se obtenga el efecto inverso: grandes explosiones de las poblaciones de herb�voros, como consecuencia de la desaparici�n de sus parasitoides por efecto de los insecticidas. Cuando cesa la aplicaci�n, las poblaciones de mariposas regresan a sus niveles habituales de baja densidad.
El segundo factor que mantiene bajas las densidades de insectos herb�voros es el clima desfavorable. Una sucesi�n de a�os malos puede hacer disminuir las poblaciones hasta niveles realmente bajos e incluso provocar su extinci�n local. Existe una escuela de ec�logos de insectos que sostiene que el principal determinante de los tama�os poblacionales en insectos es el clima.
El tercer factor que puede disminuir la abundancia de los herb�voros es la baja densidad alimenticia de algunas partes o de toda la planta. Ya sea por sequedad, bajas concentraciones de nitr�geno asimilable o presencia de una enorme gama de sustancias t�xicas, no todo lo que parece comida para un herb�voro lo es. Como adem�s los insectos tienen en general poco tiempo para escoger los sitios de oviposici�n o de alimentaci�n, y con cierta frecuencia cometen errores, resulta que sus densidades poblacionales pueden mantenerse bajas a causa de la incapacidad para encontrar los sitios adecuados en el tiempo disponible. Por ejemplo, existe una poblaci�n de cierta mariposa en los Estados Unidos que regularmente oviposita el 50% de los huevos en una planta completamente t�xica para sus larvas. La planta contiene la misma sustancia que normalmente le indica a la mariposa cu�les son sus sitios de alimentaci�n, por lo que, enga�ada por el olor, la mariposa oviposita sobre ella.
Consecuentemente, ser�a posible pensar que la mayor�a de los insectos fit�fagos
tuvieran un efecto relativamente menor sobre los par�metros demogr�ficos de
sus plantas de alimentaci�n. Sin embargo, tambi�n es muy conocido que el otro
grupo de herb�voros (los interactivos), s� afectan m�s o menos fuertemente la
demograf�a de dichas plantas. Varios efectos interesantes se derivan de la existencia
de una relaci�n interactiva entre herb�voros y plantas. En primer lugar, la
interacci�n puede regular los tama�os de las poblaciones, con las consecuencias
para el control biol�gico de malezas que analizaremos posteriormente. Por ejemplo,
un modelo con espacio de fase como el de la figura IV.7 ha sido propuesto para
explicar el gran �xito en el control de la plaga de los nopales en Australia.
Adem�s, cuando los herb�voros producen un desequilibrio en el balance demogr�fico
de las plantas que utilizan, se propicia que la estructura de las comunidades
de plantas tambi�n se altere. Por ejemplo, John Harper y sus estudiantes de
la Universidad de Gales han demostrado con mucha claridad c�mo la acci�n de
los herb�voros puede desbalancear la competencia entre especies de plantas y
alterar totalmente la composici�n de, por ejemplo, un pastizal.
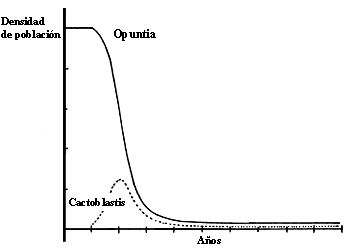
Figura IV.7. Disminuci�n de la poblaci�n de cactos por la introducci�n de
Cactoblastis cactorum, de acuerdo con un modelo matem�tico propuesto
por Lawton, J. y Caughley, en May, R. (1981) Theoretical Ecology, Blackwells
Press.
En segundo lugar, cuando el sistema es interactivo, y por lo tanto la demograf�a de ambas poblaciones se afecta, podemos esperar que resulte un proceso coevolutivo, del cual hablaremos a continuaci�n.
Alrededor de los a�os sesenta, Fraenkel propuso que los compuestos secundarios (sustancias qu�micas no directamente relacionadas con el metabolismo energ�tico, alimenticio o constructivo de las plantas) observados en las angiospermas eran utilizados por las plantas como defensas qu�micas en contra de los herb�voros. Pocos a�os despu�s, Paul Ehrlich y Peter Raven analizaron las pautas de distribuci�n de compuestos secundarios en las plantas, correlacion�ndolas con las pautas de alimentaci�n de grupos de mariposas. Concluyeron afirmando que, en primer lugar, las sustancias secundarias de las plantas desempe�aban el papel dominante en determinar las pautas de utilizaci�n de los herb�voros, y en segundo lugar, propusieron el modelo coevolutivo que se ha convertido en cl�sico: una especie de planta en la que por mutaci�n se genere un compuesto secundario que la proteja de los herb�voros, podr�a dar origen a toda una l�nea evolutiva (radiaci�n adaptativa), hasta que a su vez alguna especie de herb�voros produjera una mutaci�n que permitiera metabolizar la defensa qu�mica del grupo de plantas previamente protegido, lo cual inducir�a en los herb�voros un proceso de radiaci�n adaptativa propio. Cuando posteriormente alguna planta inventara otra defensa, se reiniciar�a el proceso. La teor�a de Ehrlich y Raven ha tenido gran importancia para el desarrollo de la ecolog�a de las interacciones planta-herb�voro: provee de hip�tesis para explicar algunos fen�menos muy interesantes, como la gran diversidad de angiospermas (cerca de trescientas mil especies), de insectos fit�fagos (trescientas cincuenta mil especies) y de compuestos secundarios (m�s de treinta mil estructuras caracterizadas), y tambi�n da una explicaci�n para el hecho de que los herb�voros no extingan a las plantas. Por otra parte, el modelo coevolutivo es generalizable a casi todas las defensas con las que cuentan las plantas (compuestos secundarios, baja calidad nutricional, cripsis o mimesis, espinas, pelos, escamas y enemigos naturales asociados). Sin embargo, algunas de estas defensas son mucho m�s sutiles que los meros venenos, y los herb�voros pueden tener problemas para contrarrestarlas, Por ejemplo, muchas plantas producen sustancias relacionadas con las hormonas que regulan la metamorfosis de los insectos. Durante la metamorfosis, los insectos se transforman m�s o menos radicalmente hasta alcanzar la fase de adulto; una larva de insecto que se alimente con ciertas plantas, como los helechos del g�nero Polypodium, puede sufrir severas aberraciones durante su metamorfosis y morir sin llegar a adulto.
Otras plantas secretan sustancias parecidas a las se�ales de alarma de algunos herb�voros. Por ejemplo, los investigadores ingleses Pickett y Gibson encontraron que una variedad de papa silvestre produce la feromona que el pulg�n Myzus persicae secreta al ser atacado por parasitoides. La planta huele entonces como si estuviera infestada de enemigos naturales del herb�voro. Es dif�cil que los herb�voros contrarresten este tipo de defensas, puesto que el compuesto secundario que la planta usa para evitar ser comida es una de las respuestas fisiol�gicas normales del insecto.
En otras ocasiones, los insectos contrarrestan sin problemas las defensas de las plantas. Por ejemplo, muchas familias de vegetales secretan l�tex pegajosos que dificultan o imposibilitan la masticaci�n de las hojas. Pues bien, como respuesta a esto, hay muchas especies de insectos que primero cortan las venas de las hojas por donde fluye el l�tex y despu�s atacan la hoja ya inerme. Otros herb�voros han llevado todav�a m�s lejos su adaptaci�n a burlar las defensas de las plantas, como aquellos que no s�lo son inmunes a los venenos de �stas, sino que los acumulan y los utilizan en su propia defensa. Tal es el caso de la mariposa Monarca, que acumula los gluc�sidos cardiacos de las plantas de las cuales se alimenta (varias especies del g�nero Asclepias), con lo que se convierte en muy poco agradable para la mayor�a de sus depredadores. Este mecanismo se puede demostrar experimentalmente criando larvas de mariposas Monarca sobre variedades de plantas que carecen o poseen los gluc�sidos cardiacos y luego alimentando a las aves que las depredan con las mariposas adultas: aqu�llas alimentadas con plantas que contienen gluc�sidos resultan inaceptables para las aves.
Cuando una especie de herb�voro encuentra una respuesta evolutiva para las defensas de una planta, se esperar�a que las presiones selectivas sobre dicha planta produjeran a la larga poblaciones con nuevas defensas. Este proceso coevolutivo se ilustra de manera clara con las diferentes adaptaciones en la "carrera armamentista" de las mariposas del g�nero Heliconius y sus plantas de alimentaci�n, las "pasionarias", del g�nero Passiflora, sistema que ha sido estudiado al detalle por Lawrence Gilbert, William Turner y Gordon Smiley.
Las pasionarias son plantas de los tr�picos americanos. Se sabe que contienen compuestos secundarios (alcaloides y glic�sidos cianog�nicos) y tiene muy pocas especies de herb�voros: pr�cticamente s�lo los Heliconius y unas pocas especies de escarabajos. Estos insectos ya han encontrado respuesta para el primer problema coevolutivo, ya que son capaces de tolerar la bater�a de compuestos secundarios de las Passifloras. M�s a�n, las mariposas los utilizan para su propia protecci�n.
Los Heliconius son los principales herb�voros y pueden tener un efecto demogr�fico severo sobre las plantas, tanto porque las hembras prefieren ovipositar cerca de las yemas de crecimiento de aqu�llas, como porque las mariposas son extremadamente eficientes para detectar a las plantas. Revisitan con regularidad a las Passifloras y oviponen sobre la misma planta, por lo que pueden defoliar completamente a un individuo. Bajo dichas circunstancias, se esperar�a que esta especie vegetal hubiese respondido evolutivamente con un nuevo conjunto de defensas. Entre �stas encontramos: 1) Enemigos naturales asociados. Las Passifloras tienen gl�ndulas que secretan n�ctar y que mantienen a hormigas y parasitoides visit�ndolas continuamente. Tanto unas como otros dan cuenta de un muy alto porcentaje de larvas o huevecillos de las mariposas. 2) Mimesis. Los Heliconius se encuentran entre las especies de mariposas con mejor visi�n, m�s alta longevidad y pautas de conducta m�s complejas. Tienen rutas fijas de b�squeda, viven hasta por seis meses, examinan visualmente las plantas antes de ovipositar y aprenden de su experiencia. Por lo tanto, no es sorprendente que las plantas respondan con adaptaciones relacionadas con estas capacidades visuales y conductuales de las mariposas. Muchas especies de Passifloras tienen formas de hojas at�picas, muy parecidas a las de plantas existentes en la zona pero inaceptables para las mariposas. Otras producen peque�as estructuras muy semejantes a huevos de Heliconius. Se ha podido demostrar experimentalmente que estos falsos huevecillos provocan un efecto disuasorio en la mariposa hembra que busca un sitio para ovipositar. Posiblemente la raz�n de esto sea que una sola planta no puede mantener simult�neamente a varias larvas, por lo que la hembra evita ovipositar sobre plantas ya ocupadas. Con sus falsos huevecillos la planta da la impresi�n de ya estar ovipositada y por lo tanto de que hay riesgo de que un nuevo huevecillo pueda ser canibalizado o no encuentre suficiente alimento. 3) Por �ltimo, existen unas pocas especies de Passifloras en las que se encuentran pelos r�gidos y agudos llamados tricomas, los cuales desgarran la delicada piel de las larvas m�s j�venes y convierten a la planta en inutilizable.
�Qui�n va ganando la carrera armamentista entre Passifloras y Heliconius? Podr�a decirse que nadie, puesto que ambos grupos de especies permanecen en el campo de batalla. Las presiones selectivas est�n ah�. Mientras los sistemas gen�ticos y morfol�gicos de los contendientes provean suficiente variabilidad heredable, deberemos esperar nuevas adaptaciones para contrarrestar las del oponente en el juego existencial.
Pese a su verosimilitud no debemos terminar este cap�tulo sin mencionar que hay muchas objeciones ecol�gicas al funcionamiento del modelo coevolutivo. El punto principal es que para que el modelo funcione, es necesario que la interacci�n se mantenga intensa y sostenida por periodos muy largos de tiempo. Dado todo lo que se sabe sobre la variabilidad clim�tica, la acci�n de los enemigos naturales y la gran heterogeneidad medioambiental, es poco probable que los herb�voros sean lo suficientemente abundantes como para mantener la pureza de la interacci�n por el lapso necesario para operar una verdadera coevoluci�n. Casos tan claros como el de Heliconius-Passiflora siguen siendo raros, ya sea porque no abunden en la naturaleza o porque los ec�logos no los hayan detectado a�n.
Recapitulando, las interacciones -+ presentan una tendencia a las oscilaciones que puede ser neutralizada por factores puramente ecol�gicos, como la heterogeneidad medioambiental, o por cambios evolutivos o coevolutivos, como la aparici�n de diversas versiones de "depredadores prudentes" (presas altamente defendidas o depredadores menos virulentos o efectivos). El papel que tienen las interacciones -+ en la estructuraci�n de las comunidades puede ser muy importante, como se demuestra por trabajos como los de Paine, Harper, etc., tanto per se como por su efecto en otras interacciones (competencia). Las presiones selectivas originadas por la interacci�n han generado adaptaciones verdaderamente espectaculares en los depredadores para obtener su alimento, as� como en las presas para evitar ser comidas, y en muchos casos el proceso selectivo se va alternando en la llamada coevoluci�n.