II. Y CRUZANDO SUPIERON
LA HERENCIA DE LOS CARACTERES DISCRETOS
SE LLAMA caracteres discretos a aquellos que pueden cambiar cualitativamente. Tal es el caso de los caracteres que us� Mendel, como la presencia o ausencia de cierta condici�n, por ejemplo tener ch�charos verdes o amarillos por un lado, o arrugados o lisos por el otro. El estudio de estas caracter�sticas, como el de todas aquellas que el genetista estudia, se lleva a cabo cruzando individuos que tienen una condici�n diferente en ellas. Por ejemplo, lo primero que a uno se le ocurrir�a para analizar la herencia de la longitud de la nariz ser�a cruzar a un individuo de nariz corta con otro de nariz larga. De esta cruza uno podr�a esperar varios tipos de resultados, pero los dos m�s diferentes ser�an los siguientes. El primero ser�a que los hijos de la cruza tuvieran una nariz intermedia entre los dos padres y el segundo ser�a que los hijos tendr�an narices intermedias, largas o cortas independientemente de la apariencia de los padres. Este segundo resultado ser�a del todo sorprendente pero significar�a que el tama�o de la nariz es independiente de los genes y que est� determinado por variables ambientales que afectan la expresi�n del genoma (el conjunto de genes).
Un ejemplo no tan extremo de un car�cter parcialmente determinado por el ambiente lo representa la altura que tenemos los seres humanos. Cu�ntas veces no hemos o�do que las generaciones recientes tienen un promedio de altura mayor que las generaciones pasadas. "Es que los j�venes de hoy tienen una mejor alimentaci�n", se dice. Por otro lado, sabemos que normalmente personas de estatura peque�a tienen hijos tambi�n de baja estatura. Lo que indicar�an estos hechos es que, en parte lo chaparro de una persona se debe a que tiene padres bajos, pero otra proporci�n se debe a la alimentaci�n que ha tenido durante su vida.. El efecto de los genes puede entonces verse oscurecido por un ambiente extremo.
Regresemos a nuestro ejemplo de las narices largas y cortas y supongamos que no existen efectos ambientales fuertes; en este caso, el efecto de los genes se expresar� (recordemos que la otra alternativa es que la progenie de la cruza de individuos con nariz corta y larga tenga una nariz de longitud intermedia). �C�mo nos resuelve esto el misterio de la herencia? De hecho no nos resuelve nada. Una alternativa que Darwin pens� al observar este tipo de resultado es que de las sangres del padre y de la madre se obten�a una mezcla intermedia. Es como si se mezclara pintura roja y blanca, el resultado ser�a pintura rosa.
Mendel resolvi� este problema al cruzar entre s� a los individuos producidos en la primera cruza. El resultado obtenido fue sorprendente pues se recuperaban las caracter�sticas existentes en los individuos de la primera generaci�n o generaci�n parental. Es decir; reaparec�an las narices largas y las cortas. pero tambi�n hab�a intermedias. Este resultado no era nuevo para el hombre ni para el genetista. En esto Mendel no aport� una novedad. Fue el m�todo de an�lisis lo que lo distingui� de sus antecesores. Encontr� que dentro de ciertos rangos de variaci�n, aproximadamente un cuarto de esta generaci�n ten�a la caracter�stica de uno de los abuelos y otro cuarto ten�a la caracter�stica alternativa (narices cortas y largas respectivamente). La otra mitad de los nietos ten�a narices intermedias.
De este an�lisis Mendel concluy� que los caracteres de los abuelos no se mezclan como si fueran pintura, sino que se mantienen independientes unos de otros. Este descubrimiento es lo que ahora se ense�a con el nombre de la primera ley de Mendel y es t�cnicamente llamada como la ley de la segregaci�n independiente de los genes. La representaci�n de este resultado con letras trajo consigo otro descubrimiento que revolucion� tambi�n nuestra concepci�n del material hereditario; para obtenerlo cada individuo necesita tener la posibilidad de poseer dos versiones alternativas de cada gene, uno heredado de su padre y otro de su madre. As�, por ejemplo, si proponemos que uno de los abuelos era A1A1 y el otro A2A2, la segregaci�n de estos alelos producir� gametos A1 en uno de los abuelos y gametos A2 en el otro, de tal manera que al unirse en la fertilizaci�n tendremos solamente hijos A1A2 que tendr�n un aspecto intermedio entre los dos originales. Esta descripci�n encierra los principios de la gen�tica que representan en la actualidad la columna vertebral de esta ciencia. Aun as�, la visi�n que Mendel ten�a de estos fen�menos no se materializaba en lo que hoy conocemos como genes, cromosomas, c�lulas, gametos y fertilizaci�n. Mendel pensaba que los caracteres consist�an en part�culas que no se mezclaban unas con otras; que al tener hijos las posibilidades gen�ticas de un organismo se divid�an en dos y, por �ltimo, que para que se originara un nuevo individuo se necesitaba la contribuci�n por partes iguales de su padre y de su madre.
Estos principios ahora son considerados muy sencillos por los hechos y conceptos generados durante este siglo, pero los principios de Mendel fueron complicados de entender para sus contempor�neos e incluso para los que leyeron su art�culo m�s de treinta a�os despu�s de que fue escrito.
Si una persona tiene ojos claros y uno de sus padres tambi�n, podemos inferir que el otro padre debe de ser heterocigoto, con un alelo de ojos claros y otro de ojos oscuros. Este ejemplo ilustra no solamente la concepci�n de dominancia sino otros conceptos que ayudaron a Mendel a entender el fen�meno de la herencia. Ya explicamos anteriormente que Mendel concibi� la idea de que los genes son particulados, es decir; que cada uno de nosotros hereda un gene de su padre y otro de su madre. Estos dos genes pueden ser iguales y entonces decimos que se trata de un homocigoto (homo significa igual, es decir; el cigoto est� formado por dos iguales). Por otro lado estos dos genes pueden ser diferentes y entonces se dice que el individuo heterocigoto (hetero significa diferente). En individuos que son homocigotos la expresi�n de los genes no presenta ning�n problema porque los dos genes son iguales. Si uno de ellos lleva la orden de hacer ojos claros y el otro tambi�n, se har�n ojos claros. Pero, �qu� pasa cuando los dos genes son diferentes? En general, en este caso, se puede obtener uno de dos resultados. El primero es que los dos genes se expresen y la apariencia de estos heterocigotos sea intermedia entre ambos. As�, por ejemplo, de la expresi�n de un gene que produce semillas amarillas y otro que las produce sin color tendr�amos semillas amarillas claras. La otra posibilidad es que uno de los dos genes se exprese y el otro permanezca sin expresarse. Esto suceder�a en el caso de los ojos que ya mencionamos, ya que si se mezcla un gene para ojos claros con otro para ojos oscuros, resultar�a un individuo de ojos oscuros. En este caso se dice que uno de los genes (el que s� se expresa) domina sobre la expresi�n del otro.
Mendel trabaj� con ch�charos que presentan caracter�sticas con este tipo de dominancia. Entre estos dos extremos, sin dominancia y con dominancia completa hay variantes intermedias, de tal manera que �stos son s�lo extremos de un continuo. En el caso de las semillas amarillas, por ejemplo, el individuo heterocigoto se parecer�a m�s a las semillas amarillas pero no ser�a tan oscuro. Este concepto nos permite entonces dividir el grado de dominancia en tres tipos: la dominancia total, la dominancia parcial y la ausencia de dominancia. Esta �ltima define a los genes como codominantes. Un ejemplo de estas relaciones entre los alelos (que son las diferentes opciones que tiene un gene) es el de los tipos de sangre O, A y B. En este sistema existen tres posibles alelos que son precisamente el alelo O (a veces tambi�n llamado cero), el alelo A y el alelo B. En este sistema los alelos A y B son codominantes entre s� pero dominan al O. As�, por ejemplo, si tenemos un individuo con dos alelos diferentes A y B, el tipo de sangre ser� AB pero si cualquiera de estos alelos se combina en otro individuo con el O, entonces el tipo de sangre ser� o A o B seg�n el caso. La �nica manera de tener un tipo de sangre O es teniendo dos alelos O en un individuo. Si entonces alguien nos dice que su tipo de sangre es B, puede tener una combinaci�n de alelos (un genotipo) BO o BB pero no podemos estar seguros porque la dominancia enmascara el genotipo.
De estos ejemplos podemos entonces concluir lo que se ha dado en llamar el principio de la segregaci�n de los caracteres que no es m�s que la expresi�n de que cada uno de los progenitores genera dos tipos de alelos; �stos pueden ser iguales o diferentes pero se separan uno del otro en una forma cualitativa, es decir; se segregan.
La segunda ley de Mendel: La herencia independiente de caracteres
Pero, �qu� pas� cuando Mendel cruz� individuos que difer�an ya no en un solo car�cter sino en dos? En la tabla 1 se pueden observar los resultados que se obtuvieron al cruzar dos variedades de toloache (Datura stramonium) de frutos prickly y flores rojas con toloache, con frutos lisos y flores blancas en la segunda generaci�n.
TABLA 1. Resultados obtenidos de la cruza entre el toloache de frutos
prickly y flores rojas, con toloache de frutos lisos y flores blancas
| Primera generación: | Todas las plantas presentaron frutos prikly |
| y flores rojas | |
| Segunda generación: | 204 toloaches con frutos prikly y flores rojas |
| 65 toloaches con frutos prikly y flores blancas | |
| 81 toloaches con frutos lisos y flores rojas | |
| 13 toloaches con frutos lisos y flores blancas |
De esta tabla se desprenden varios datos importantes. En primer lugar; si analizamos los dos caracteres utilizados por separado encontramos las relaciones num�ricas predichas por Mendel al hablar de un solo car�cter. As�, por ejemplo, se obtuvieron en la segunda generaci�n 269 plantas con frutos prickly y 94 con frutos lisos, y los valores que Mendel esperar�a ser�an 272.25 y 90.75, respectivamente. Por otro lado, se obtuvieron 285 plantas con flores rojas y 78 con flores blancas, cuando los resultados esperados por Mendel en este caso ser�an tambi�n 272.25 y 90.75 respectivamente. Estas relaciones son esperadas porque si suponemos segregaci�n independiente de los alelos y dominancia completa en ambos tipos de caracteres por parte de los caracteres prickly (P) y rojo (R) entonces la primera generaci�n ser� de genotipo PpRt; donde las min�sculas representan los alelos que no son dominantes (recesivos) y que en nuestro caso ser�an el fruto liso y las flores blancas. En consecuencia, la segunda generaci�n se obtendr� de la uni�n de cuatro tipos de gametos, PR, Pr; pR y pr; que existiendo en frecuencias iguales se unir�n formando diecis�is genotipos en la siguiente forma:
| PR | Pr | pR | pr | |
| PR | PPRR PR, RO |
PPRr PR, RO |
PpRR PR, RO |
PpRr PR, RO |
| Pr | PPRr PR, RO |
PPrr PR, BL |
PpRr PR, RO |
Pprr PR, BL |
| pR | PpRR PR, RO |
PpRr PR, RO |
ppRR LI, RO |
ppRr LI, RO |
| pr | PpRr PR, RO |
Pprr PR, BL |
ppRr LI, RO |
pprr LI, BL |
Si contamos el n�mero de plantas con fenotipo prickly y liso encontramos que ser�n 12 y 4, respectivamente. Esta misma relaci�n se aplica al color de la flor. Encontraremos, 12 plantas con flores rojas por cada cuatro de color blanco. Ésta es una relaci�n de tres a uno y la que obtenemos a nivel de fenotipo (apariencia), pero no de genotipo. La raz�n de ello es que al existir la dominancia los genotipos RR y Rr tendr�n la misma apariencia, as� como aquellos PP y Pp. Adem�s en este caso se puede obtener las relaciones esperadas de los fenotipos considerando dos caracter�sticas. De cada 16 plantas se esperan nueve prickly de flor roja, 3 con fruto prickle y flor blanca, tres de fruto liso y flor roja y una de fruto liso y flor blanca. Lo que estos datos nos indican es que las suposiciones que llevamos a cabo para generar los gametos y los individuos son ciertas siempre y cuando los datos observados no sean diferentes de aquellos esperados. La primera suposici�n fue que para cada caracter�stica los alelos se segregan uno de otro, como lo dice la primera ley de Mendel. La segunda suposici�n fue que la combinaci�n de alelos de diferentes caracter�sticas es independiente del alelo que se trate, es decir; existe la misma probabilidad de que se formen gametos con los alelos PR con que los Pr; y de hecho, supusimos que la proporci�n de cada uno de los gametos ser�a de un cuarto tanto en el polen como en las c�lulas femeninas. Esta es la segunda ley de Mendel y se refiere a la formaci�n de h�bridos utilizando dos caracter�sticas. Esto lo describi� Mendel dentro del cap�tulo de cruzas dih�bridas y fue la parte menos entendida tanto por sus contempor�neos como por al menos dos de sus redescubridores (De Vries y Tschermak). Aparentemente fue Correns el �nico que comprendi� el significado de esta ley.
El ejemplo del toloache lo tomamos no solamente por el significado medicinal que esta planta tiene y ha tenido dentro de la cultura mexicana, sino tambi�n porque fue una especie utilizada por otro genetista que, aunque no es reconocido como uno de los redescubridores de las leyes de Mendel, desempe�� un papel muy importante en su difusi�n a principios de este siglo. Me refiero a William Bateson, quien por muchos a�os represent� en Inglaterra el mendelismo m�s profundo, como veremos un poco m�s adelante. Como evidencia de que Bateson ya estaba a punto de redescubrir el mendelismo se pueden ofrecer algunos hechos. Por ejemplo, en 1899 en la Conferencia Internacional de Hibridizaci�n, Bateson present� los datos que acabamos de ver para el toloache sin reconocer en ellos la regularidad num�rica que Mendel propon�a. Aun as�, al terminar su presentaci�n afirm� que mediante el an�lisis estad�stico de la progenie de la hibridizaci�n entre diferentes individuos se podr�a entender m�s el significado de sus resultados. Adem�s se refiri� al an�lisis de caracter�sticas individuales. De hecho, desde 1894 Bateson llam� a la comunidad cient�fica a enfrascarse en experimentos de hibridizaci�n que aunque requieren de mucho esfuerzo y recursos, ayudan "a empezar a saber". Bateson reconoci� desde muy pronto que al cruzar individuos con caracter�sticas diferentes el fen�meno de la herencia podr�a empezar a ser comprendido.
La asociaci�n de los genes y los coromosomas
Durante los primeros a�os de este siglo hab�a dos escuelas gen�ticas contendiendo por la supremac�a de los principios que rigen la herencia de caracter�sticas: la escuela biometrista y la mendelista. Aunque este tema lo tocaremos m�s adelante, aqu� habr�a que decir que el verdadero desarrollo postmendeliano se consolid� con la intervenci�n de un grupo de genetistas que trabajaban con la mosca de la fruta (Drosophila melanogaster). Ellos eran Thomas H. Morgan, Alfred H. Sturtevant, Calvin B. Bridges (1889-1939) y Hermann J. Muller. Antes que ellos hubo dos investigadores que sentaron las bases para que se generara la teor�a cromos�mica de la herencia, W. S. Sutton y Th. Boveri.
A finales del siglo pasado ya se hab�an descrito los cromosomas. Estos cuerpos que aparec�an durante la meiosis y la mitosis y desaparec�an en las otras fases celulares planteaban fuertes interrogantes acerca de su funci�n. Fueron Sutton y Boven quienes primero reconocieron no s�lo la individualidad de los cromosomas sino tambi�n los identificaron como los portadores de los genes. Esta individualidad cromos�mica significaba que pod�an reconocerse distintos cromosomas una y otra vez en distintas mitosis, pero la prueba m�s importante que apoy� esta idea fue un experimento de Boven en el que se demostr� que se requer�a de los 36 cromosomas en una especie de erizo para formar un individuo normal, lo que suger�a adem�s que los cromosomas se complementaban en una forma muy particular y que por ello no conten�an informaci�n redundante. Sutton, por otro lado, demostr� que para una especie de saltamontes los machos ten�an siempre un cromosoma menos que las hembras, de tal manera que los factores o genes que determinan el sexo en estos animales est�n localizados en los cromosomas. Estos dos descubrimientos generaron las bases de lo que se llam� posteriormente la teor�a cromos�mica de la herencia que en su forma m�s completa fue propuesta por Morgan, Bridges, Muller y Sturtevant.
El caso de la mosca de ojos blancos
Hacia 1909 Morgan empez� a trabajar con la mosca de la fruta. Estos animales
tienen com�nmente ojos rojos, pero un d�a en el laboratorio se encontr� un macho
que ten�a ojos blancos. Este descubrimiento sugiri� una serie de preguntas que
el grupo de Morgan empez� a tratar de responder muy r�pidamente. �De d�nde apareci�
este individuo? �Al cruzarlo con hembras de ojos rojos qu� tipo de moscas se
producir�an? �Se comportar�a este car�cter en forma estrictamente mendeliana?
Sin duda lo que se deb�a de hacer primero era cruzarlo para empezar a responder
estas preguntas. Los resultados de estas cruzas fueron sorprendentes ya que
no se cumpl�an en todos los casos las leyes de Mendel. Al cruzar al macho de
ojos blancos con sus hermanas de ojos rojos, Morgan encontr� que toda la progenie
ten�a ojos rojos sugiriendo que el car�cter era recesivo pero sorpresivamente
en la siguiente generaci�n reaparecieron machos de ojos blancos pero ninguna
hembra de ojos rojos. Morgan y sus colegas razonaron que si el gene del color
de ojos estaba localizado en el cromosoma que determina el sexo se pod�an explicar
todos sus resultados utilizando para ello solamente las leyes de Mendel (Figura
4).
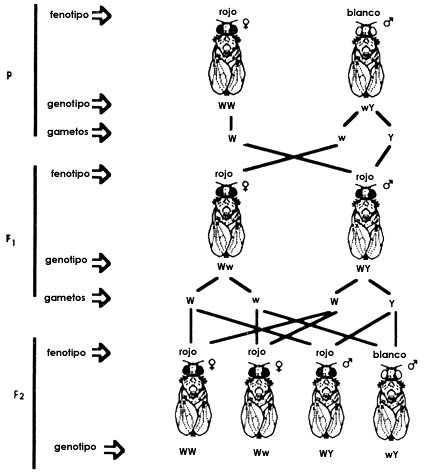
Figura 4. Herencia de la mutaci�n que transforma en blanco el color normal
del ojo (rojo) de la mosca de la fruta Drosophila Melanogaster.
La determinaci�n del sexo en diferentes animales tiene varios mecanismos. La
determinaci�n del sexo de la Drosophila es parecida a la que se hace
en humanos, pero no es igual. Mientras que en la mosca de la fruta la mayor�a
de los machos tienen el cromosoma sexual y, todos los hombres tienen un cromosoma
Y en sus c�lulas. Las hembras, entonces, adem�s de tener un cierto n�mero
de cromosomas que no determinan el sexo (tres pares en Drosophila, 23
en el hombre) tienen dos cromosomas sexuales llamados X. Se dice
que en estos casos los machos son el sexo heterogam�tico porque producen gametos
diferentes, unos con un cromosoma X y otros con uno Y.
Las hembras son el sexo homogam�tico ya que producen s�lo gametos con cromosomas
X, adem�s de los otros que no determinan el sexo. La diferencia
entre Drosophila y el hombre es que mientras en el hombre el cromosoma
Y determina que el cigoto sea macho, en Drosophila es el
n�mero de cromosomas X lo que determina el sexo del producto; si
se tiene una relaci�n de uno a uno o m�s entre los cromosomas X
y los cromosomas que no determinan el sexo del producto ser� una hembra aunque
exista un cromosoma Y, mientras que si la relaci�n es cercana a
un medio el producto ser� un macho. As� por ejemplo un individuo XXY
en Drosophila es una hembra, pero en el hombre es un macho. En los dos
casos es un complemento anormal de cromosomas y produce individuos con problemas
gen�ticos, pero estos casos extraordinarios nos ayudan a entender el mecanismo
por medio del cual se determina el sexo. Este fen�meno se pod�a ver en el microscopio
a finales del siglo pasado y a principios de �ste. Los cit�logos (que estudian
las c�lulas) se dieron cuenta de que en especies como la mosca de la fruta los
machos generan dos tipos de gametos con cromosomas que se ven distintos, mientras
que en las hembras todos los gametos son iguales. Este hecho permiti� distinguir
por medio del microscopio a los cromosomas sexuales y por otro lado, usar marcadores
gen�ticos para seguir a los genes. Por ejemplo, en la Drosophila los
machos con ojos blancos heredaban estos alelos s�lo a las hembras, mientras
que hembras de ojos blancos (homocigotas recesivas) heredaban estos alelos tanto
a sus hijas como a sus hijos. As� (y en contra de lo esperado por Mendel), de
una cruza entre hembras de ojos blancos y machos de ojos rojos se producen machos
de ojos blancos y hembras de ojos rojos. En un gene mendeliano cl�sico, y dado
que rojo domina sobre el blanco, se obtendr�a una progenie con ojos rojos. Esto
se debe a que los machos s�lo tienen un alelo para este gene y �ste es heredado
de su madre. De su padre no heredan alelos para esta caracter�stica. Las hembras,
por otro lado, s� se comportan normalmente y heredan un alelo de su padre y
otro de su madre. En la figura 5(a) se representa este tipo de cruza, adem�s
de los procesos que ocurren en la meiosis y que apoyan f�sicamente este tipo
de comportamiento gen�tico. Los cromosomas, como lo expresa la primera ley de
Mendel, segregan uno de otro y los genes segregan en la misma manera. La herencia
de ambos sigue las mismas reglas y por ello debemos esperar que exista una asociaci�n
f�sica entre los cromosomas y los genes. De aqu� se concluye entonces que las
part�culas hereditarias descritas por Mendel y que posteriormente se llamaron
genes est�n localizados en los cromosomas.
La no disyunci�n de los cromosomas: una evidencia m�s de la teor�a cromos�mica de la herencia
En la cruza de machos de Drosophila de ojos rojos con hembras de ojos
blancos uno espera y obtiene, seg�n hemos descrito hasta ahora, que todos los
machos producidos tengan ojos blancos ya que reciben su cromosoma X de
sus madres, y que todas las hembras los tengan rojos porque reciben un cromosoma
X de su padre y otro de su madre. Con una frecuencia muy baja (de
uno en 2 000), sin embargo, se obtienen machos de ojos rojos y hembras de ojos
blancos, lo cual no parecer�a tener una explicaci�n clara, seg�n lo que hemos
visto hasta ahora. Bridges, uno de los colaboradores de Morgan, propuso que
lo que ocurrir�a en estos casos extraordinarios era que durante la meiosis los
cromosomas X de la madre no se separaban en dos c�lulas diferentes,
es decir; no hab�a la disyunci�n que se espera normalmente (Figura 5(a). En
la figura 5(b) se puede apreciar que de la uni�n de los gametos anormales de
la hembra (uno con dos cromosomas X y el otro sin cromosomas) con
los gametos normales de los machos se obtienen dos individuos que normalmente
mueren, uno de ellos con sólo un cromosoma Y y el otro con
tres cromosomas X. Los otros dos, un macho con ojos rojos y con
s�lo un cromosoma X y una hembra de ojos blancos y con dos cromosomas
X y uno Y son los individuos que Morgan y sus colegas
consideraban raros. Este fen�meno, una vez más, permiti� asociar el comportamiento
de los cromosomas y el de los genes, comprobando con evidencia independiente
que los genes est�n f�sicamente localizados en los cromosomas. Este fen�meno,
por otro lado, no es espec�fico de Drosophila, varias enfermedades humanas
se explican por este hecho, la no disyunci�n de los cromosomas durante la meiosis.
Un ejemplo conocido es el s�ndrome de Down y los individuos que lo padecen tienen
un total de 47 cromosomas debido a la presencia de tres cromosomas en lo que
se llama el par 21 (los cromosomas humanos son nombrados del par 1 al 22, aparte
del par de cromosomas sexuales, en funci�n de su tama�o, de tal manera que el
par 21 es uno de los pares con cromosomas mas peque�os). Estos descubrimientos
demostraron sin lugar a dudas la relaci�n f�sica que existe entre los genes
y los cromosomas, siendo �stos los portadores de aqu�llos.
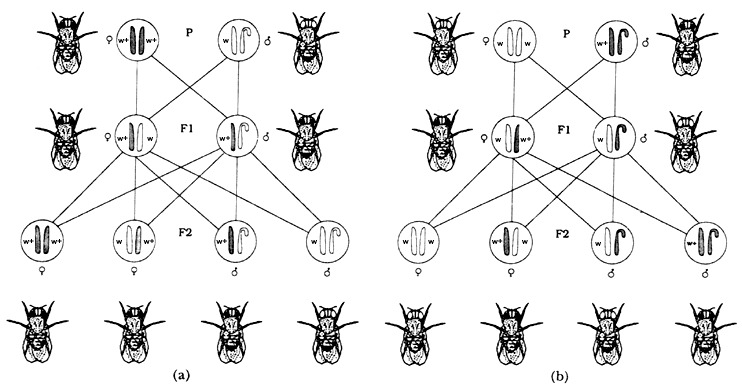
Figura 5. En (a) vemos un caso de la herencia ligada al sexo en la Drosophila
melanogaster cruzando una hembra de ojos rojos con un macho de ojos blancos
(w+y w). En (b) otro caso de herencia ligada al sexo cruzando una hembra de
ojos blancos con un macho de ojos rojos. Podemos notar que ambos resultados
son diferentes.
Pero la teor�a cromos�mica de la herencia no solamente predec�a que los genes
estaban en los cromosomas sino tambi�n que aquellos genes que est�n en el mismo
cromosoma no cumplir�n la segunda ley de Mendel, es decir la uni�n independiente
de alelos de diferentes caracteres. Como se demostr� posteriormente los siete
caracteres utilizados por Mendel en sus cruzas con ch�charos est�n localizados
en cada uno de los siete pares de cromosomas de los ch�charos, de tal manera
que en ellos s� se cumple la predicci�n de que los alelos se van a comportar
como si fueran f�sicamente independientes unos de otros. De hecho, los resultados
del grupo de Morgan acerca de la asociaci�n f�sica entre los genes y los cromosomas
se generaron al descubrir que, como ya hemos visto, el car�cter que determina
el sexo en Drosophila no se asociaba de manera independientemente con
el gene que determina el color de los ojos. Es decir, la demostraci�n de que
los genes est�n en los cromosomas llev� a la conclusi�n te�rica de que en algunas
ocasiones, cuando los genes considerados se encuentran en el mismo cromosoma,
no se cumple la segunda ley de Mendel. Me parece que �ste es un buen ejemplo
de c�mo la respuesta a una pregunta importante trae como consecuencia necesaria
otras preguntas. Al existir una relaci�n f�sica entre algunos de los genes es
de esperar que, al formarse los gametos, los alelos que pertenecen a genes en
el mismo cromosoma tender�n a mantenerse en los mismos gametos. La figura 6
muestra qu� ocurrir�a si �ste fuera el caso. En la primera generaci�n se producen
individuos heterocigotos para los dos genes y los gametos que se producen de
estos individuos ser�n AB y ab, si no hay ninguna de recombinaci�n. A ellos
se a�adir�an los gametos Ab y aB si hubiera uni�n independiente de caracteres.
Cuando los genes se encuentran en cromosomas diferentes entonces la proporci�n
de cada uno de los cuatro gametos mencionados ser� de un cuarto, pero si se
encuentran en el mismo cromosoma las proporciones depender�n del grado de recombinaci�n
que exista entre ellos.
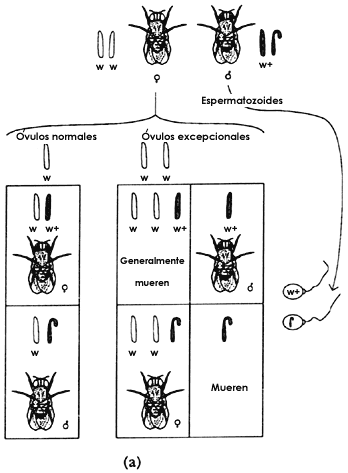
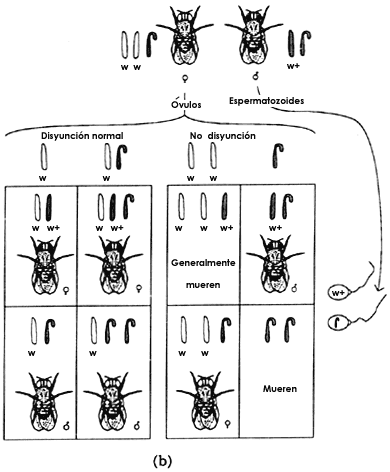
Figura 6. (a) La no disyunci�n primaria en la Drosophila melanogaster
con los resultados normales. (b) Bridges propuso la hip�tesis de que la segregaci�n
en la meiosis de las hembras con dos cromosomas X y uno Y
puede efectuarse en dos formas: 1) los dos cromosomas X se desplazan
a polos diferentes, uno de ellos acompa�ado de un Y, lo que sucede
en 96% de los casos, y 2) los cromosomas X se van a un polo y el
Y al otro 4% restante de los casos.
La recombinaci�n gen�tica y su evidencia física
Durante la meiosis (Figura 7) los cromosomas del padre y de la madre se aparean
para posteriormente repartirse en las dos c�lulas que formar�n los gametos.
Durante ese apareamiento los cromosomas intercambian informaci�n entre ellos
de tal manera que se forman cromosomas que son una combinaci�n de los cromosomas
del padre y de la madre. Si el cromosoma paterno tiene en su cromosoma los alelos
A y B en dos genes distintos y el materno tiene los alelos a y b en esos mismos
genes y ocurre una de recombinaci�n entre estos genes, entonces los cromosomas
que ir�n a formar las c�lulas hijas tendr�n complementos gen�ticos que no exist�an
en sus padres, como son las combinaciones Ab y aB. Estos gametos se llaman gametos
recombinantes, en contraposici�n a los gametos ab y AB que se llaman parentales
por ser los que fueron heredados de los dos padres. Esta recombinaci�n entre
dos genes depende en gran parte de la distancia f�sica a la que los dos genes
est�n en el cromosoma, de tal manera que la frecuencia con la que ocurren es
una medida de la distancia f�sica entre los genes considerados. La figura 8
muestra un mapa de los siete cromosomas del ch�charo, y fue construido utilizando
la metodolog�a de mapeo gen�tico, as� como una cruza de dos genes que codifican
para enzimas, la cual se emple� para obtener la distancia gen�tica entre ellos.
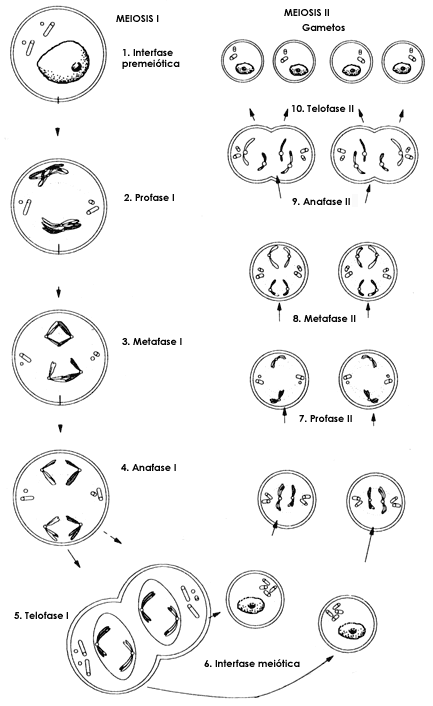
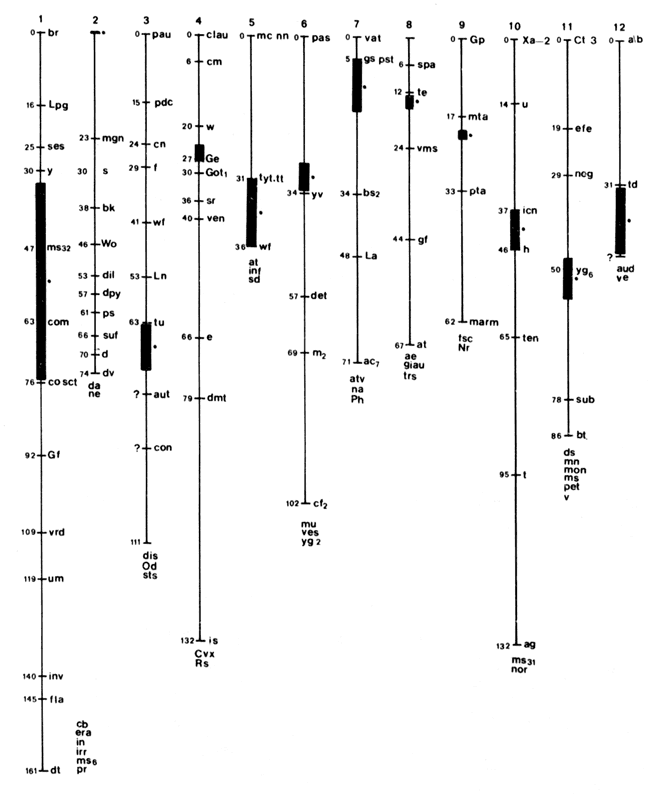
Figura 8. Mapa cromos�mico del tomate.
A principios d� este siglo se observ� c�mo durante la meiosis los cromosomas se entrelazan para formar lo que se llama quiasmas. Posteriormente se demostr� que este entrelazamiento significaba un intercambio de material cromos�mico entre la madre y el padre. Fue a Sturtevant, quien entonces ten�a dieciocho a�os, al que se le ocurri� que deber�a haber una correlaci�n entre la probabilidad de recombinaci�n y la distancia entre dos genes. De hecho, en 1913, junto con Morgan, Sturtevant public� el primer mapa gen�tico del cromosoma sexual X de la Drosophila. Una vez m�s se unieron las evidencias recabadas por los genetistas (la recombinaci�n) y los cit�logos (los quiasmas) para agregar otro punto a la teor�a cromos�mica de la herencia.
La distancia gen�tica no siempre es igual a la distancia f�sica en los cromosomas
No son estructuras sencillas los cromosomas de los organismos como Drosophila o el hombre, que tienen c�lulas con un n�cleo (eucariontes, a diferencia de aquellos organismos que no tienen n�cleo como por ejemplo las bacterias, y que pertenecen al grupo de los procariontes). Simplemente, el hecho de que sean observables con un microscopio �ptico se demuestra que los cromosomas est�n formados por mol�culas sencillas. Al hacer un an�lisis de la composici�n qu�mica de los cromosomas se encontr� que est�n compuestos por dos tipos de mol�culas: prote�nas y �cidos nucleicos. Las prote�nas se encuentran en la parte externa de los �cidos nucleicos. Se ha encontrado que en algunas partes de los cromosomas las prote�nas est�n en una proporci�n mayor que en otras y en esas zonas es menor la frecuencia de quiasmas, y consecuentemente la de recombinaci�n. Este fen�meno, la falta de homogeneidad de los cromosomas a lo largo de toda su longitud, hace que la distancia gen�tica no siempre corresponda con la distancia f�sica y por ejemplo habr� zonas en donde la frecuencia de recombinaci�n sea relativamente alta, pero en donde f�sicamente los genes est�n muy cercanos unos de otros y viceversa.
No todos los genes de los organismos con n�cleo est�n en el n�cleo
Hasta ahora hemos considerado que los genes est�n localizados en los cromosomas que se encuentran en el n�cleo, que nuestro complemento cromos�mico es la uni�n entre la mitad de los cromosomas de nuestro padre y la mitad de los de nuestra madre. En ese contexto las leyes de Mendel han funcionado bien, menos en el caso en el que dos genes est�n f�sicamente ligados en un mismo cromosoma. Sin embargo, existen otras excepciones a las leyes de Mendel; por ejemplo, en algunos casos se ha encontrado que tanto en plantas como en animales algunas caracter�sticas son transmitidas por uno solo de los sexos (com�nmente la madre). Es decir; en estas caracter�sticas un individuo hereda el genotipo y la apariencia de la madre y no del padre. Posteriormente, se ha encontrado que organelos celulares que est�n en el citoplasma, como la mitocondria y el cloroplasto, contienen �cido desoxirribonucleico (ADN) que se replica independientemente de aquel que se encuentra en el n�cleo. Asimismo, se ha encontrado que este tipo de organelos contienen toda la maquinaria para llevar a cabo la s�ntesis de prote�nas y lo que es m�s, esta maquinaria es m�s parecida a la de aquellos organismos sin n�cleo que a la de aquellos que tienen n�cleo. Estos datos, unidos con otros de naturaleza similar sugieren que estos organelos son el resultado de una antigua simbiosis entre organismos procariontes. De aquella simbiosis tan estrecha se han generado organismos m�s complejos en los que es ya dif�cil distinguir las barreras entre las dos especies originales. Para entender la estructura de los genes que se encuentran en organelos, como los cloroplastos y las mitocondrias, analizaremos la de los organismos procariontes.
La gen�tica de los organismos sin n�cleo
Los organismos procariontes, como ya hemos visto, tienen una organizaci�n gen�tica m�s sencilla que los eucariontes. Sus cromosomas no son estructuras complejas y consisten casi solamente de mol�culas de ADN. Estos cromosomas son circulares, a diferencia de los cromosomas eucariontes que tienen una mol�cula lineal de ADN. Una de las caracter�sticas m�s importantes es que los organismos procariontes son haploides; para reproducirse, una c�lula se divide a la mitad dando lugar a dos c�lulas hijas. Previamente, el ADN se duplica de tal manera que a cada c�lula le corresponde una copia completa del genoma de la c�lula parental. Este sistema de reproducci�n impide en principio que se puedan llevar a cabo cruzas como las que hizo Mendel con los ch�charos. Por esto, para los organismos procariontes se utilizan otros m�todos de an�lisis gen�tico.
Al igual que en los eucariontes, los m�todos de mapeo gen�tico se basan en la existencia de la recombinaci�n entre dos genomas que contienen informaci�n gen�tica diferente. El problema en estos organismos es que por ser haploides, no es frecuente que tengan dos genomas f�sicamente cerca para llevar a cabo la recombinaci�n gen�tica. Esta recombinaci�n es similar a aquella que Morgan y sus colaboradores describieron para Drosophila. Despu�s de que dos mol�culas de ADN se aparean por tener secuencias hom�logas (que contienen informaci�n muy parecida), se rompen en uno o varios puntos y se unen ya no con la hebra de la cual se rompieron sino con otra que contiene los mismos genes pero distintos alelos. Esto hace que se junten alelos de diferentes genes que estaban separados. El genetista usa esta informaci�n para evaluar la distancia entre los genes considerados. La ventaja de utilizar organismos procariontes es que por ser haploide la mayor�a de los fenotipos representa a los genotipos que los producen sin que exista la dominancia.
Otro aspecto caracter�stico de los procariontes consiste en las caracter�sticas que se analizan. Estas no pueden ser; como en el caso de los ch�charos de Mendel, lo rugoso de la semilla o el color de la vaina. En las bacterias, las caracter�sticas que com�nmente se prefieren son las capacidades para crecer en medios que no contienen diferentes compuestos, como son los amino�cidos o las vitaminas. Normalmente las bacterias tienen la capacidad de sintetizar la mayor parte de los amino�cidos necesarios para hacer prote�nas, por ejemplo, a partir de unos pocos compuestos, pero en algunos casos, y por tener defectuoso el mecanismo de s�ntesis de amino�cidos, para crecer en un medio requieren m�nimamente de la adici�n del amino�cido del cual perdieron la capacidad de sintetizar. As�, por ejemplo, se dice que una cepa es aux�trofa para leucina si perdi� la capacidad de producir leucina. Normalmente, �stos son los fenotipos que el genetista de bacterias analiza y, como se puede ver, estos fenotipos no son estrictamente iguales a los que se analizan en plantas o animales multicelulares. Aun as�, el concepto de fenotipo mantiene su significado original ya que es la expresi�n de una condici�n gen�tica particular que en este caso es la habilidad de biosintetizar un compuesto en especial.
En el caso de los virus el an�lisis gen�tico es tambi�n diferente, ya que la cantidad de informaci�n gen�tica que tienen es mucho menor. Al no ser organismos multicelulares no tienen los fenotipos que Mendel analiz� en los ch�charos, pero tampoco tienen sistemas biosint�ticos ya que utilizan los sistemas de la c�lula hospedera, de tal manera que tampoco se utilizan caracter�sticas relacionadas con la capacidad de crecer o no en medios con diversos compuestos. En los virus, entonces, lo que com�nmente se ha tomado en cuenta para llevar a cabo an�lisis gen�ticos es su habilidad de infectar a esta o aquella variedad o cepa de bacteria o el aspecto que tiene el lisado (ruptura) de las c�lulas bacterianas. Nos referimos especialmente a c�lulas bacterianas porque son las que com�nmente se han utilizado, pero no debemos olvidar que los hospederos que utilizan diferentes tipos de virus incluyen tambi�n c�lulas eucariontes y no solamente c�lulas procariontes. Para observar las caracter�sticas gen�ticas del virus invasor en bacterias lo que se hace es poner a la bacteria en una gelatina que contiene los elementos m�nimos para su crecimiento. Una vez que la poblaci�n de bacterias forma una capa de crecimiento continua sobre la gelatina se a�ade el virus que infecta a las bacterias, crece en ellas y despu�s de un tiempo las rompe. Cuando este fen�meno ha ocurrido en la mayor parte de las c�lulas bacterianas la colonia cambia su consistencia. Este es un aspecto en el cual difieren los virus. Entonces, el an�lisis de la gen�tica de virus y bacterias no se realiza como en el caso de los ch�charos de Mendel, por el an�lisis de los individuos parentales y de la progenie de �stos, pues tienen una tasa de crecimiento tan alta que se requiere multiplicarlos durante varias generaciones para observar el resultado del experimento gen�tico. Otro aspecto que merece ser mencionado es que los virus contienen una cantidad muy peque�a de genes, de tal manera que un cambio en casi cualquiera de ellos imposibilitar�a al virus para infectar a las bacterias. As� es imposible analizar la gen�tica de un virus que ni siquiera puede crecer y que no lo podernos ver. Para analizar estos genes letales (porque provocan la muerte de su portador;) se estudia la manera en que los cambios de temperatura hacen que a veces sí puedan infectar y a veces no (digamos a 40°C).
Utilizando un sistema en el que crecen los mismos virus a dos temperaturas diferentes se pueden detectar aquellos que se desarrollan a la temperatura permisiva (digamos 30°C). El cambio que han experimentado estos virus en su material gen�tico es de tal naturaleza que les impide infectar a su hospedero.
M�TODOS DE MAPEO DE GENES EN PROCARIONTES
Cuando un genetista trabaja con un eucarionte (como Mendel trabaj� con ch�charos)
sigue un procedimiento m�s o menos sencillo para detectar si dos alelos corresponden
a un mismo gene. Por ejemplo, si una caracter�stica afecta el color de los ojos
o de una semilla, uno puede estar m�s o menos seguro de que mapear�n en la misma
posici�n dentro de un cromosoma. Haciendo cruzas se puede determinar si los
alelos segregan en forma mendeliana, y por lo tanto si son alelos de un mismo
gene. Dado que en virus y bacterias �ste no es el caso, porque el fenotipo que
se analiza es la habilidad de infectar cierta cepa de bacterias o la de no crecer
en un medio donde falta un amino�cido, en primer t�rmino tenemos que asegurar
que los alelos pertenecen a un mismo gene. Para ello, lo que normalmente se
hace es estudiar si los alelos se complementan o no. As�, por ejemplo, si a
cierta cepa de bacteria inyectamos simult�neamente dos virus que de manera independiente
no pueden infectar y se produce una infecci�n, lo que esto querr� decir es que
las dos mutaciones mapean en genes diferentes. Si por otro lado encontramos
que no hay infecci�n, entonces podremos asegurar que las dos mutaciones pertenecen
a un mismo gene y que por lo tanto son alelos de �l (Figura 9(a)). Despu�s de
determinar si las dos mutaciones no se localizan dentro del mismo gene se puede
proceder a mapearlos, es decir, a determinar la distancia gen�tica que hay entre
ellos. Esto se lleva a cabo tomando en cuenta el mismo principio que se usa
en eucariontes, es decir se utiliza la fracci�n de los individuos producidos
que presentan el fenotipo recombinante. Es importante mencionar que, a diferencia
de la complementaci�n, esta fracci�n tiene un valor mucho m�s peque�o, de tal
manera que es muy sencillo distinguir entre ambas. En la figura 9(b) se muestra
un ejemplo de los resultados obtenidos con un virus de bacterias que ha sido
muy utilizado en estudios gen�ticos.
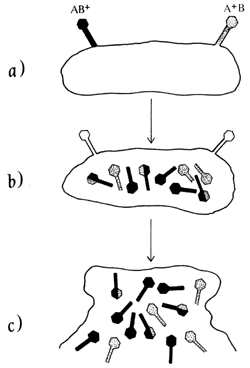
Figura 9 (a). Cruza de fagos mutantes. a y b que se logra
infectando al hospedero en condiciones que permitan la replicaci�n de los fagos.
En a se da la infecci�n simult�nea y en b la proliferaci�n. En
c la progenie es estudiada en condiciones permisivas para determinar
el n�mero total de todos los genotipos posibles: AB+, AB, A+B,
y A+B+. Tambi�n se estudia la progenie en condiciones restrictivas para determinar
el n�mero de genotipos A+ B+.
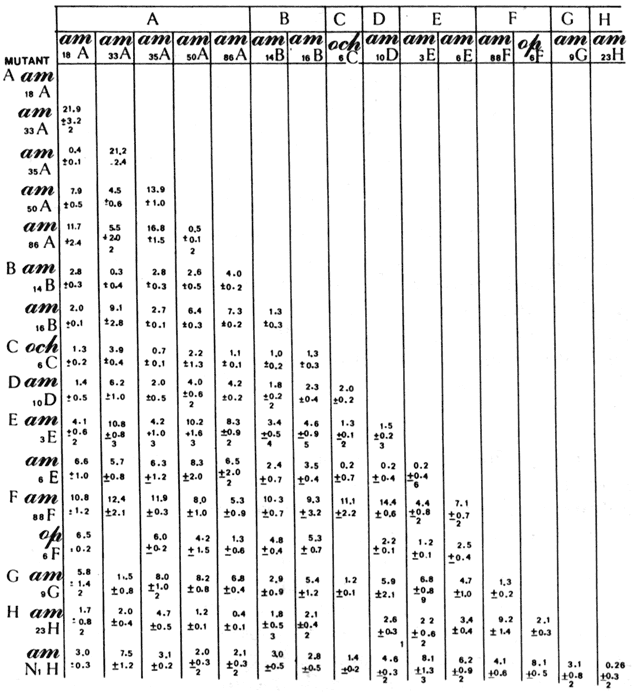
Figura 9(b). En la tabla podemos observar las frecuencias de recombinaci�n en cruzas mutantes ø X 174.
En este ejemplo se ve que los mutantes am l8 y am35 no complementan, lo cual indica que est�n en el mismo gene, mientras que los mutantes am l8 y am 88 complementan, por lo que se considera que est�n en diferentes genes. Aplicando este tipo de an�lisis a una gran cantidad de mutantes de este virus se ha podido determinar que hay nueve grupos de complementaci�n, tambi�n llamados cistrones o genes, que afectan diferentes funciones y prote�nas de este virus. Este tipo de an�lisis se llev� a cabo entre 1965 y 1975 y posteriormente se ha podido seguir la secuencia de todo el genoma de este virus utilizando m�todos de secuenciaci�n de ADN, que han demostrado que los genes est�n dispuestos en el orden predicho en los an�lisis gen�ticos, pero adem�s este organismo es de los pocos que han sido secuenciados completamente en su ADN, que uno de los cistrones est� anidado dentro de otro, de tal manera que al menos en este caso no se cumple el modelo propuesto por Morgan, seg�n el cual los genes est�n arreglados en los cromosomas como las cuentas de un rosario, uno despu�s de otro.
Los procedimientos que se han utilizado para llevar a cabo el mapeo de genes en las bacterias se basan tambi�n en la recombinaci�n y se ha aprovechado la existencia de mecanismos de transferencia de material gen�tico de una bacteria a otra. Estos procesos incluyen dos m�todos diferentes. El primero es la conjugaci�n y el segundo la transducci�n. Como veremos más adelante, en esta segunda interviene en forma directa un tipo de virus particular que tiene la habilidad de insertarse dentro del genoma de la bacteria.
La conjugaci�n bacteriana consiste en la transferencia de material gen�tico de una c�lula a otra; una vez que el material se ha transferido se incorpora al genoma de la c�lula receptora en forma de un pl�smido que tiene la misma estructura de un cromosoma, pero m�s peque�o. La manera en que se transfieren genes localizados en el cromosoma de la bacteria es llevando a cabo una recombinaci�n entre el pl�smido y el cromosoma, de tal manera que al ponerse en contacto una bacteria con otra el pl�smido incorporado al cromosoma lleva consigo algunos de los genes que hab�a en el cromosoma. Adem�s, una vez que los genes est�n dentro de la c�lula receptora pueden recombinarse con aquellos del cromosoma, y en funci�n de las caracter�sticas que codifican los genes analizados se selecciona en un medio de cultivo aquellos que representan los recombinantes que se estima como una fracci�n del total de colonias crecidas, y por lo tanto es posible calcular la distancia gen�tica entre cada uno de los marcadores.
La transducci�n bacteriana se lleva a cabo en dos pasos principales. El primero es la infecci�n de una bacteria por un virus. En muchos casos despu�s de infectar a las c�lulas los virus se reproducen utilizando el mecanismo de sus v�ctimas, hasta que llega un momento en el que son tantos que hacen que la c�lula bacteriana se rompa y los libere al exterior. En otros casos, al invadir la c�lula los virus no toman control de los mecanismos de metabolismo celular sino que se insertan en el cromosoma bacteriano, donde permanecen por algunas generaciones celulares hasta que por factores del medio ambiente el virus se separa del cromosoma y se comporta como un virus infeccioso, rompiendo la c�lula bacteriana. Hasta aqu� toda esta descripci�n no parece tener que ver con mecanismos de mapeo de genes, pero esto adquiere otra perspectiva si consideramos que en la mayor�a de las ocasiones, cuando el virus del cromosoma se separa lleva consigo pedazos del cromosoma bacteriano que var�an en su tama�o y que por lo tanto pueden contener genes bacterianos que nosotros quisi�ramos mapear. Cuando este virus infecta otra bacteria, y considerando que contiene genes hom�logos a ella, se puede llevar a cabo una recombinaci�n de tal manera que los genes de la bacteria original y los de la bacteria final se intercambiar�n a una frecuencia que depender� nuevamente de la distancia f�sica entre ellos y con esa frecuencia podremos localizar las posiciones relativas de los genes en la misma forma que se hace para organismos eucariontes.
LA HERENCIA DE LOS CARACTERES CUANTITATIVOS
Los caracteres que Mendel analiz� eran caracteres de calidad, o sea que se distingu�an uno de otro por aspectos de apariencia cualitativos. Tal era el caso de los ch�charos lisos y rugosos o verdes y amarillos. Existe otro tipo de caracter�sticas que son cuantitativas, como por ejemplo el peso o la altura. A finales del siglo pasado no se sab�a si los caracteres cuantitativos obedec�an las leyes de Mendel o no. Francis Galton (1822-1911), primo de Charles Darwin, fue uno de los primeros en enfrentar este problema con t�cnicas estad�sticas que en ese tiempo se empezaban a utilizar. La caracter�stica que utiliz� fue la altura de las personas. Razon� que si la altura se heredaba de padres a hijos, deber�a existir una relaci�n lineal entre el promedio de la altura de los padres y aquella de los hijos. Esta relaci�n (Figura 10) se corrigi� en dos aspectos principalmente. El primero de ellos es la observaci�n de que com�nmente las mujeres son m�s bajas que los hombres, de tal manera que sus datos se ajustaron suponiendo que su altura es, en promedio, el 80% de aquella de los hombres. La segunda correcci�n se refiere a la expresi�n de los resultados como una desviaci�n, ya sea positiva o negativa, de la media poblacional. Existe una clara relaci�n entre estas variables que Galton interpret� como una evidencia fuerte de que existe un componente gen�tico de la altura en las poblaciones humanas. De hecho, Galton, a partir de este resultado, propuso una teor�a gen�tica de la herencia, de caracteres cuantitativos, diferente de los principios propuestos por Mendel. El enfoque de Galton estaba basado en una concepci�n estad�stica de la herencia, pero a finales del siglo pasado apenas se estaban desarrollando los estudios estad�sticos de la relaci�n entre dos variables. Posteriormente se demostr� que este enfoque, si se analizaba en un contexto gen�tico, ten�a muchas posibilidades de desarrollo; se vio que por ejemplo, exist�a una interpretaci�n gen�tica clara de los par�metros de la relaci�n propuesta por Galton. Estos resultados se obtuvieron 20 a�os despu�s y para que esto ocurriera se tuvo que desarrollar una de las mayores controversias de que la gen�tica tiene memoria. Esta controversia se desarroll� entre los seguidores de Galton que posteriormente se llamaron biometristas y aquellos que apoyaban la idea de que las leyes de Mendel eran universales. Como ocurre en la mayor�a de las controversias cient�ficas, en algunos aspectos una de las teor�as tiene raz�n mientras que en otros es la teor�a alternativa la m�s atinada. Lo importante de esta situaci�n es que se plantean preguntas concretas que tratan de ser respondidas por una gran cantidad de investigadores o al menos m�s r�pidamente que otras preguntas que no generan controversias).
La esencia de la diferencia entre las escuelas era que seg�n una de ellas,
la mendeliana, todas las caracter�sticas segu�an las leyes de Mendel, mientras
que para la otra, las caracter�sticas cuantitativas (que determinan gran parte
de la adaptaci�n de los organismos) obedecen a otros principios que, como los
propuestos por Galton, se derivaban de principios estad�sticos. Estas �ltimas
caracter�sticas, dec�an los biometristas, eran las que realmente importaban
para la evoluci�n, y no si un ch�charo era liso o rugoso.
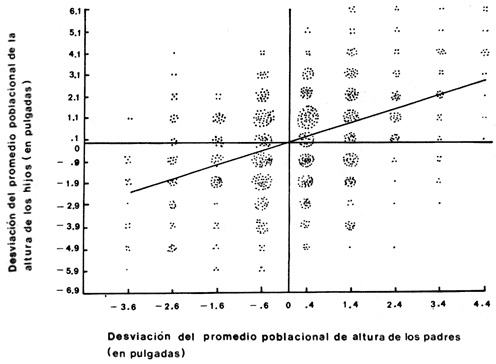
Figura 10. La figura muestra en contraste la altura de los hijos contra la altura promedio de los padres. La pendiente de la regresi�n es la heredabilidad, h� y su valor en este caso es de 0.65.
Herencia dura y herencia blanda
Antes de continuar con la controversia biometrista mendelista es importante se�alar que antes de que fueran redescubiertas las leyes de Mendel, la teor�a de la evoluci�n de Charles Darwin (1809-1882) consideraba que hab�a dos tipos de herencia. La primera de ellas se deb�a a la condici�n del organismo, como dir�amos ahora, a sus genes; y la segunda a mecanismos del ambiente que de alguna manera modificaban la herencia de los organismos y facilitaban que se heredara a las siguientes generaciones. Este segundo mecanismo era el que originalmente hab�a propuesto Jean Baptiste Lamarck (1741-1829) y que Darwin consideraba aplicable a ciertas caracter�sticas. Durante todo el final del siglo pasado los bi�logos y genetistas se debat�an entre estas alternativas hasta que August Weismann (1834-1914) marc� el inicio del descr�dito de la herencia de los caracteres adquiridos. Para hacerlo se bas� en tres evidencias: 1) en los estudios citol�gicos que realiz� no encontr� una forma que explicara f�sicamente el efecto del medio sobre la l�nea germinal; 2) todos los casos en los que se mencionaba la herencia de caracteres adquiridos se pod�an explicar por medio de la selecci�n natural darwinista, y 3) en insectos sociales como hormigas y termitas no se pod�a explicar la forma en que evolucionaban las caracter�sticas de las castas que no siguen un modelo de herencia de caracter�sticas adquiridas al reproducirse.
Hemos tra�do a colaci�n a Weismann porque en la controversia que estamos analizando renaci�, como ha ocurrido regularmente, la importancia del ambiente en la determinaci�n del fenotipo. Los exponentes iniciales de esta controversia fueron William Bateson (1861-1926), mendelista, quien en funci�n del conocimiento de la �poca ten�a una concepci�n tipol�gica de la especie, as� como la idea de que la evoluci�n se mueve a saltos y no en una forma gradual, como hab�a propuesto Darwin. En el otro lado de la controversia se encontraba Walter E. R. Weldon (1860-1906), quien era de la escuela de Galton y por ello ten�a una concepci�n estad�stica del proceso de la herencia. La controversia lleg� a tener una gran cantidad de consideraciones personales y despu�s de varias cartas y publicaciones, sobre todo en la revista Nature, Bateson y Weldon dejaron de hablarse, de tal forma que los que finalmente resolvieron la controversia fueron investigadores que no estaban mezclados en ella en un principio.
El primer genetista que arroj� luz sobre este problema fue W. Johanssen (1857-1927), quien trabaj� con frijoles para demostrar que la apariencia de un individuo (el fenotipo) tiene un componente gen�tico (el genotipo) y un componente ambiental que no es heredable. Esto lo demostr� con experimentos de selecci�n artificial en familias (frijoles derivados de una sola planta, la madre) que ten�an en promedio diferente peso de las semillas, pero que dentro de una familia de frijoles sujetos a autofertilizaci�n constante (es decir; que el polen de un individuo se utiliza para fertilizar el estigma del mismo individuo) hab�a variabilidad en el peso de las semillas producidas. La causa principal de esto eran aspectos ambientales, ya que en el proceso de autofertilizaci�n la familia se va haciendo cada vez m�s homocigota hasta que despu�s de seis a diez generaciones se puede considerar una l�nea pura gen�ticamente.
Una vez demostrado el hecho de que una porci�n del fenotipo se debe a factores ambientales y otra a factores en g�neticos, se llevaron acabo algunos experimentos para estimar estas proporciones. En Estados Unidos, Edward M. East trabaj� con la especie Nicotiana longiflora de la familia del tabaco, y como Johanssen, obtuvo l�neas puras generadas de la continua autofecundaci�n de plantas seleccionadas por tener la corola grande o peque�a. Despu�s de varias generaciones de autofertilizaci�n East midi� la variaci�n del tama�o de la corola en las l�neas puras y la interpret� como el componente ambiental del fenotipo. East posteriormente cruz� a dos individuos representativos de los tama�os de corola, el peque�o (de aproximadamente 4 cm de largo) y el mayor (de alrededor de 10 cm). Los individuos resultantes de la cruza ya no ser�an homocigotos pero seguir�an siendo iguales entre s�, con un alelo heredado de su madre y el otro de su padre. En esta generaci�n, al igual que en la anterior, toda la variabilidad encontrada entre el tama�o de la corola se debe a factores ambientales y no a factores gen�ticos, ya que todos los individuos tanto de la l�nea materna, de la paterna y de la primera generaci�n filial son id�nticos entre sí gen�ticamente. Pero qu� pasa en la segunda generaci�n filial si cruzamos entre s� a los individuos heterocigotos. Siguiendo las leyes de Mendel lo que ocurrir� es que los alelos de los diferentes genes segregar�n lo cual dar� lugar a una gran cantidad de genotipos que cubrir�n tanto el extremo de flor peque�a como el de la flor m�s grande. As� en esta segunda generaci�n filial encontraremos que la variabilidad presente tendr� causas gen�ticas y ambientales. De esta manera, y si restamos la variaci�n fenot�pica total, el promedio de la variaci�n encontrada en la primera generaci�n filial y la generaci�n parental la obtendremos haciendo un promedio (Figura 11), tendremos as� la variabilidad gen�tica. As�, al comparar estos datos podremos definir la proporci�n de la variabilidad fenot�pica que se debe a factores gen�ticos y a factores ambientales.
En el experimento anterior supusimos que la herencia de los caracteres cuantitativos
segu�a las leyes de Mendel y de hecho la controversia entre biometristas y mendelistas,
se resolvi� a favor de los mendelistas pero hubo otro experimento clave que
ayud� a definir la situaci�n. Este lo hizo el genetista sueco Herman Nilsson-Ehle
(1873-1949), quien trabaj� con trigo. Nilsson-Ehle utiliz� l�neas puras que
ten�an el color de la semilla desde blanca hasta un color rojo oscuro. Al cruzar
las blancas con semillas rojas oscuras obtuvo una primera generaci�n intermedia
como se esperar�a en un car�cter mendeliano determinado por un solo gene, pero
en la siguiente generaci�n, en vez de tener una cuarta parte de los hijos con
semillas blancas o rojo oscuras, de cada 64 semillas obtuvo una de color blanco
y una de color rojo oscuro. Estos resultados se pueden explicar siempre y cuando
supongamos que son tres y no uno los genes que determinan el color de la semilla
y que en cada uno de los genes hay dos alelos (Figura 11). De esa manera se
pueden obtener siete diferentes clases de color, desde un blanco total hasta
un rojo oscuro, dependiendo del n�mero de alelos para el color que tenga la
semilla.
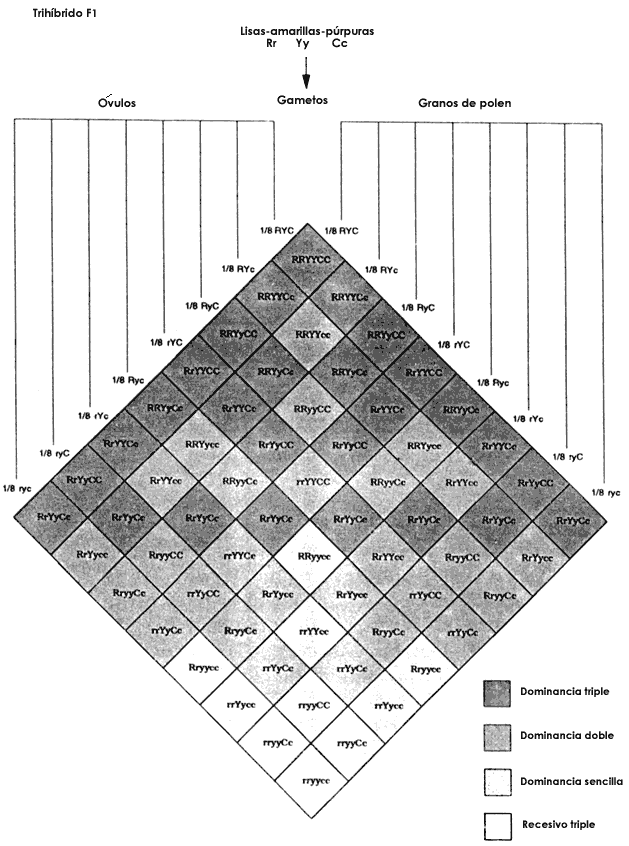
Figura 11. Genotipos producidos en la progenie de individuos trih�bridos
autofecundados o cruzados entre s�. Hay 64 combinaciones de 8 gametos paternos
y 8 gametos maternos, pero que corresponden s�lo a 27 genotipos diferentes.
Si hay dominancia, como en el ejemplo mostrado, los 27 genotipos se reducen
a 8 fenotipos. El ejemplo presentado corresponde al de los ch�charos de Mendel
trih�bridos para liso, rugoso, amarillo y verde y p�rpura y blanco.
Con los experimentos de East, Johanssen y Nilsson-Ehle se pudo definir que la variaci�n fenot�pica se descompone en una parte gen�tica y en una ambiental (Johanssen), que estas porciones se pueden estimar usando por ejemplo el enfoque de East y que en casos en los que la variaci�n es continua las leyes de Mendel se cumplen (Nilsson-Ehle). Estos tres investigadores ayudaron a demostrar que las leyes de Mendel son universales (algo que Mendel no hab�a podido lograr) lo cual consolid� los cimientos de la gen�tica alrededor de 1915-1920. En 1918, Ronald A. Fisher (1860 -1962) escribi� un art�culo en el cual demostr� que las caracter�sticas cuantitativas tienen una explicaci�n mendeliana desde un punto de vista te�rico y no solamente emp�rico.
La historia de la gen�tica nos muestra que el concepto de gene (como la unidad de la herencia) ha variado. En un principio Mendel hablaba de ciertos deterninadores gen�ticos que causaban que una planta tuviera una caracter�stica (semilla lisa) u otra (semilla rugosa). Con el desarrollo de la gen�tica mendeliana y gracias a la teor�a cromos�mica de la herencia pudo separarse al elemento gen�tico (genes) de la caracter�stica propiamente dicha. Despu�s de varias d�cadas de trabajos experimentales se lleg� a la conclusi�n de que los genes produc�an efectos espec�ficos en los organismos. As�, ya durante la segunda d�cada del presente siglo se hab�an localizado genes que formaban parte de los cromosomas presentes en los n�cleos de las c�lulas eucariontes. Gracias a la replicaci�n de estos cromosomas y a la divisi�n celular tenemos garantizada la permanencia de los organismos vivientes. En esos momentos se cambia al de gene como la unidad de la herencia.
M�s adelante, al surgir nuevas t�cnicas se logr� conocer c�mo variaban estos genes y cu�les eran las causas.
El gene como la unidad de variaci�n
Al estudiar la naturaleza de los genes se estableci� que tienen dos propiedades fundamentalmente: primera, su capacidad de autorreplicarse y de autocat�lisis, y segunda, su capacidad de mutar, es decir; de cambiar. Esta segunda caracter�stica de los genes fue explorada hacia 1915 por uno de los estudiantes de Morgan, que pertenec�a a "el grupo de las moscas". Estos estudios y sus resultados se convirtieron en una parte fundamental de la ahora ya gen�tica cl�sica.
Estos estudios de mutag�nesis, generaci�n de variaciones, se iniciaron hacia la segunda d�cada de este siglo por H. J. Muller, en la Drosophila melanogaster. Gracias al descubrimiento de los diversos colores de los ojos de esta mosca de la fruta se pudo seguir c�mo se heredaban estas diferencias generaci�n tras generaci�n. Posteriormente, Muller descubri� que si se aplicaba radiaci�n a estas moscas antes de aparearse, se presentaban en la descendencia aberraciones o cambios no s�lo en la coloraci�n de los ojos sino en otras caracter�sticas, como la forma de las alas, la presencia de pelos en el t�rax, etc.
Muller concluy� que estos cambios eran debidos a su vez a cambios f�sicos en los genes por la acci�n de la radiaci�n. Estudios posteriores dieron la raz�n a Muller. La mutaci�n o cambio podr�a inducirse con los rayos X para despu�s estudiar c�mo se transmit�a a la descendencia. Este descubrimiento inaugur� un campo dentro de la gen�tica mendeliana que colocaba a la mutaci�n en el centro de la investigaci�n. Gracias a esto Muller recibir�a en Premio Nobel en 1947.
Una vez iniciado este campo, dentro de la gen�tica cl�sica se desarroll� el estudio f�sico de los genes. Ya no bastaba con saber c�mo eran transmitidos mendelianamente, ahora tambi�n interesaba de qu� estaban hechos y c�mo variaban. A este campo se le design� posteriormente como biolog�a molecular y se inici� con muchos trabajos encaminados a determinar c�mo est�n hechos los genes, qu� producen y c�mo lo hacen y, tambi�n, c�mo se generan las variaciones. Obviamente, esta diversificaci�n de la gen�tica en gen�tica mendeliana por un lado, y biolog�a molecular por el otro, enriqueci� nuestro conocimiento de los genes y de su funcionamiento, y tambi�n la concepci�n que se ten�a a principios del siglo XX acerca del material gen�tico.
El gene como la unidad de funci�n
Durante la �poca del florecimiento de la gen�tica mendeliana el gene era considerado como la unidad de la herencia, pero poco se sab�a acerca de c�mo funcionaba. Los genes s�lo pod�an identificarse por cambios o mutaciones que produc�an alteraciones visibles en los organismos; estos cambios pod�an ser sencillos, como la alteraci�n en el color de los ojos, o muy complejos. No fue sino hasta que la bioqu�mica se interes� en la acci�n de los genes que se modific� este concepto de gene como unidad de la herencia.
Una de las consecuencias fue el descubrimiento de que los genes producen prote�nas o, dicho de otra manera, el producto de la actividad de los genes es la formaci�n de prote�nas encargadas de m�ltiples actividades dentro de la c�lula. Estas prote�nas son las causantes de que un organismo presente una caracter�stica determinada. Por ejemplo, el color de los ojos de la mosca de la fruta, el albinismo en humanos, etc., son alteraciones gen�ticas que indican que el gene original o normal ha sido alterado y por lo tanto su producto. Este descubrimiento fue hecho por Beadle y Tatum en 1941. Ellos designaron a este trabajo como una genuina prote�na, puntualizando que la actividad de cada gene estaba indicada por la presencia de una prote�na espec�fica. En este momento, la idea del gene estaba ligada a su funcionamiento: el gene tiene una funci�n y �sta es la formaci�n de una prote�na. En la actualidad y gracias al desarrollo de t�cnicas novedosas, se sabe que no necesariamente cada gene produce una prote�na y tambi�n que no toda prote�na est� codificada por un solo gene. Es decir, existen algunas prote�nas que necesitan de la participaci�n de varios genes, y algunos genes participan en la formaci�n de m�s de una prote�na.
Vemos una vez m�s c�mo el avance de la biolog�a permiti� caracterizar de manera m�s profunda la problem�tica del gene.
En el siguiente cap�tulo hablaremos de la estructura molecular del material gen�tico y de su variaci�n.