VIII. OPERACIÓN DE LA MAQUINARIA: REGULACIÓN CELULAR
CUANDO las células llevan a cabo actividades de motilidad, un
requisito indispensable es la capacidad de movilizar iones de Ca2+
(Figura VIII.1). Como vimos en otros capítulos, para la contracción muscular
se requiere la movilización del Ca2+ de reservorios especiales al
citoplasma. La presencia del Ca2+ y su asociación a proteínas, como
las troponinas, induce varios cambios en la conformación de las proteínas del
sarcómero, favoreciendo la fosforilación de la miosina y su interacción con
la actina. En el caso de células no musculares, el Ca2+ sale también
de reservorios especiales (vesículas generalmente) hacia el citoplasma, en donde
activa a otras proteínas que al unir una o más moléculas de calcio cambian su
conformación y pueden interaccionar a su vez con otras proteínas para que éstas
funcionen. Este es el caso de la gelsolina, que se analizó en relación con el
movimiento amiboideo, y la interacción de calmodulina con microtúbulos y otras
proteínas en la mitosis. Muchas proteínas del citoesqueleto son fosforiladas
por enzimas llamadas cinasas. Cuando esto sucede su estructura se modifica,
al igual que sus relaciones con otras proteínas, dando como resultado la reorganización
de los componentes del citoesqueleto, según las necesidades de la célula. La
desfosforilación se lleva a cabo por otras proteínas presentes en todas las
células, llamadas fosfatasas. Estas enzimas se encargan de remover el o los
grupos fosfatos añadidos por las cinasas.
Como ejemplos de movimientos celulares en los que la fosforilación y desfosforilación
de proteínas es uno de los mecanismos de control tenemos a la fagocitosis, la
desorganización de la envoltura nuclear y la condensación de la cromatina que
ocurren durante la mitosis, la exocitosis de varios tipos de vesículas, la ciclosis,
la formación de contactos focales sobre el sustrato para facilitar el desplazamiento
de las células y otros movimientos que describiremos brevemente a continuación.
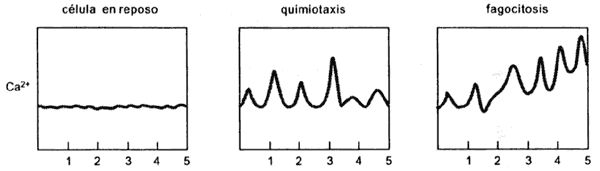
Figura VIII.1. Modificaciones del Ca2+ intracelular durante diferentes
tipos de movimiento. Los números denotan unidades de tiempo.
El desplazamiento dirigido de una célula puede ser favorecido por la presencia
de estímulos químicos que se encuentran solubles en el medio o que forman la
parte sólida del sustrato sobre el que se mueve la célula. Este fenómeno se
conoce como quimiotaxis y está bien estudiado en las bacterias que se desplazan
hacia el atrayente por rotación de su o sus flagelos. Los flagelos bacterianos
están constituidos por una proteína diferente de las tubulinas pero que también
puede polimerizarse y despolimerizarse. Las bacterias tienen receptores membranales
que reciben la señal quimio atrayente y pasan señales al interior de la célula.
La señal llega entonces al motor que mueve al o los flagelos y se produce un
cambio en la dirección de la rotación que permite a la bacteria dirigirse hacia
el estímulo. Un primer paso en la activación de señales es la fosforización
de una proteína cinasa que depende de ATP. Esta proteína puede
entonces fosforilar a otras que, ya modificadas, intervienen con el motor flagelar
y producen la rotación del flagelo. Cabe aclarar que no pueden rotar las mutantes
bacterianas que carecen de esta proteína. Al desfosforilarse la proteína se
revierte el proceso y el flagelo rota en la dirección contraria, causando un
cambio en la dirección del movimiento. Un caso bien estudiado en las células
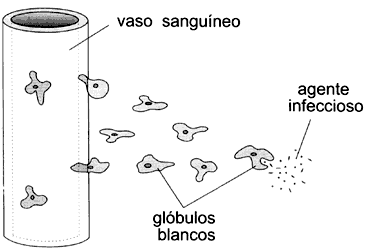
eucariontes es el de los leucocitos que son atraídos hacia un sitio de infección,
y que al ser activados modifican su forma y su velocidad de movimiento (Figura
VIII.2). Las amibas de Dictyostelium reaccionan en forma similar al estímulo
producido por AMP cíclico. Una vez cerca del estímulo las amibas
se agregan y empiezan a formar un sincicio, de donde se diferenciarán las células
sexuales. En estos casos, como en el de las bacterias, la fosforilación de las
proteínas, como respuesta al estímulo, es el efecto inicial a partir del cual
se dan los cambios de la motilidad.
Un caso particular es la estimulación de la locomoción y la secreción de proteasas,
que en varias células son producidas por las proteínas de la matriz extracelular,
como la fibronectina, la laminina y la colágena. El reconocimiento del estímulo
se hace cuando la célula, a través de una molécula proteica de su superficie
(receptor), reconoce a la molécula que es el estimulador (ligando). Cuando esto
sucede, el receptor y el ligando unidos causan una cascada de cambios en la
organización de la membraña celular que son interpretados como señales para
que en el interior de la célula haya cambios a muchos niveles. Tal vez el más
notable, en el tema que nos interesa en este libro, es la reorganización del
citoesqueleto que tiene consecuencias en la forma de la célula, en su capacidad
de desplazarse, en la velocidad con que se desplaza y en actividades metabólicas
que le permiten secretar enzimas al exterior, fagocitar e incluso dividirse
(Figura VIII.3).
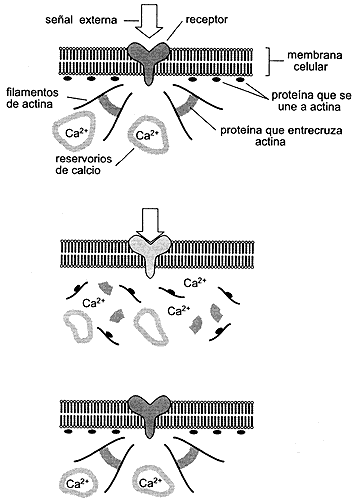
Figura VIII.3. Reorganización del citoesqueleto en una célula que corresponde
a una señal externa a través de un receptor en la membrana celular.
Los fenómenos de motilidad son muy diversos y algunas células presentan una organización especial del citoesqueleto que le permite movimientos a veces espectaculares. Esto es particularmente notable en los organismos unicelulares que se han tenido que adaptar a todos los medios, desarrollando estructuras que les permiten sobrevivir en condiciones adversas e incluso invadir a otras células como parásitos. Como es imposible hacer una revisión completa de todas las modalidades, nos hemos concentrado en ilustrar las bases generales de la organización molecular y estructural del citoesqueleto, en una célula, y considerando el movimiento individual. Desde luego, en muchos casos, el propósito de estos movimientos se hace en el contexto de un órgano o un organismo completo, formado por muchas células y actuando en concierto. El movimiento es, entonces, el resultado de la actividad del citoesqueleto de cada una de las células que forman un organismo, y sus diversas manifestaciones dependerán de la información genética de aquél y de la posibilidad de expresión de ciertos genes. La información genética se transforma, como hemos visto en este libro, en estructuras físicas con capacidad mecánica y fuerza motora de las que dependen todos los movimientos de la maquinaria celular.