VI. APARATOS DE PRECISIÓN: MOTILIDAD COORDINADA
HEMOS mencionado que el transporte intracelular cumple con funciones
muy diversas y ayuda para la adecuada distribución de metabolitos en distintas
zonas del citoplasma, para el mantenimiento de regiones muy distantes del cuerpo
celular y para el intercambio de mensajes químicos entre células relacionadas.
Todos estos procesos son desde luego invisibles; sólo pueden observarse con
la ayuda de un microscopio o de técnicas especiales y, en general, ocurren en
la profundidad de los tejidos dentro de los organismos. Existe, sin embargo,
un famoso ejemplo de transporte intracelular con efectos macroscópicos, que
se manifiesta en la parte externa de algunos animales, por lo que es fácilmente
observable, aunque en ocasiones persigue precisamente lo opuesto, es decir;
conferir al individuo que lo posee el preciado don de la invisibilidad. La evolución
ha perfeccionado este dispositivo como un eficaz medio de comunicación silenciosa
entre congéneres, o como un recurso de seguridad para la oportuna desaparición
ante los ojos de los enemigos naturales o de los distraídos prospectos para
un buen almuerzo. Dicho dispositivo se encuentra en diversos grupos zoológicos,
desde los insectos hasta los vertebrados, pero sus beneficiarios mejor conocidos
son los peces y, por antonomasia, el camaleón. Mediante modificaciones del color
que exhiben en diversas partes de sus cuerpos, muchos de estos animales pueden
declarar su pasión por un determinado ejemplar del sexo opuesto, mostrar su
ira ante un rival en amores o en dominio de territorio, o tan sólo remedar el
patrón cromático del fondo hasta fundirse ópticamente con el ambiente. Todos
estos cambios de apariencia son posibles gracias a células superficiales llamadas
cromatóforos, es decir, que transportan pigmentos.
Un cromatóforo típico consiste en una célula discoidal o en forma de estrella
aplanada con puntas ramificadas, que contiene en su interior —aparte del
núcleo, mitocondrias y demás organelos citoplásmicos— cientos e incluso
miles de gránulos de alguna sustancia colorida (Figura VI.1). Los cromatóforos
reciben el nombre específico de melanóforos si la sustancia es negra, eritróforos
si es rojiza, iridóforos si es iridiscente, etc. En respuesta a ciertos estímulos
nerviosos u hormonales, el cromatóforo acumula las partículas de pigmento en
su región central o las dispersa hasta que quedan distribuidas uniformemente
por todo el volumen celular. Como resultado de estas movilizaciones radiales
en masa, el citoplasma varía sus propiedades de absorción y reflexión de luz;
la mayor parte de su extensión palidece cuando las partículas están agregadas,
y adquiere la coloración respectiva cuando el pigmento se dispersa. Dada que
en cada escama o porción de piel ellos responden de igual manera a los estímulos,
ese sector modificará la intensidad del color correspondiente; la activación
síncrona de series de sectores contiguos cambiará así el aspecto de vastas zonas
corporales. En ocasiones un mismo sector puede contener cromatóforos de diferentes
colores —melanóforos intercalados con eritróforos y con iridóforos—
y cada tipo es controlado por estímulos distintos; el matiz cromático se verá
entonces alterado en función de la estimulación relativa de cada tipo, según
las circunstancias. La multiplicidad de combinaciones puede llegar a mimetizar
el entorno inmediato con una fidelidad sorprendente.
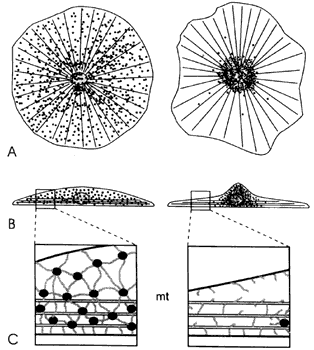
Figura VI.1. Esquema ilustrativo del movimiento de los gránulos de pigmento
dentro de un cromatóforo. A) Célula vista desde arriba; B) vista lateral; C)
sección amplificada de la célula en B), donde se ve la asociación de los gránulos
con elementos del citoesqueleto (microtúbulos mt y matriz cotoplasmica).
Como en el caso de las neuronas, la morfología peculiar de los cromatóforos
se debe a una organización especial del citoesqueleto. Hay numerosos microtúbulos
orientados con la misma polaridad desde un sitio central hacia todos los rincones
de la periferia, como los rayos de una rueda de bicicleta. Esta disposición
radial es apta para dirigir y brindar soporte a los desplazamientos de los gránulos
de pigmento, cuya migración centrífuga durante la dispersión es mediada por
la cinesina, al menos en los melanóforos. Al igual que otras manifestaciones
de transporte intracelular basado en microtúbulos, los cambios de posición del
pigmento son muy sensibles a drogas como la colchicina y la vinblastina, y dependen
estrictamente de un generoso aporte de ATP. Es evidente, por tanto,
que el mecanismo de los cromatóforos es similar al del transporte axoplásmico,
pero adaptado para funcionar en un arreglo radial más que longitudinal (Figura
VI.2).

Figura VI. 2. Micrografía que muestra tres estados de distribución de gránulos
de pigmento en cromatóforos del cangrejo de río. De izq. a der. Dispersión máxima,
dispersión media y agregación máxima del pigmento.(Fotografía cortesía de
L. Rodríguez Sosa).
Pero en los cromatóforos interviene además otro elemento del citoesqueleto, que desempeña un papel clave para aumentar en mucho la eficiencia y la coordinación del sistema. Tanto en el transporte axoplásmico como en las corrientes citoplásmicas de las células vegetales, los motores moleculares corren sueltos o asociados con partículas libres; cada componente desplazado obedece al trabajo de motores particulares con los que está directamente uncido, o al flujo general provocado por la continua marcha unidireccional de una. multitud de motores sueltos. En ambos casos se observa que las estructuras se trasladan de manera más o menos simultánea, pero independiente. En contraste, los cromatóforos muestran un movimiento concertado de los gránulos de pigmento (Figura Vl.2). Ya sea durante la agregación o durante la dispersión, cada gránulo mantiene su posición relativa con respecto al resto de la población; se dice que el movimiento es coherente. El secreto de esta uniformidad estriba en que los gránulos son transportados en un vehículo común. Las partículas de pigmento se encuentran engarzadas en un material fibroso llamado red microtrabecular. La actividad de las moléculas de cinesina, alejándose del centro del cromatóforo cuando migran sobre los microtúbulos obliga a que esta red se extienda por el citoplasma y distribuya así el pigmento en todo el volumen celular; y cuando, ante un cambio de condiciones, los motores cesan de actuar, la red se contrae por su propia elasticidad arrastrando con ella los gránulos hacia el centro de la célula. Esto provoca una mayor eficiencia de transporte, en comparación con otros sistemas, porque la acción motriz de las moléculas de cinesina es cooperativa y se transmite por medio de la red microtrabecular a muchas particulas a la vez, aun cuando no todas estén en proximidad inmediata de los microtúbulos. Al mismo tiempo se consigue una buena coordinación en los movimientos de una vasta multitud, lo que es especialmente conveniente para la función del cromatóforo. La incorporación de la red microtrabecular como un medio de transmisión de fuerzas representa así un incremento favorable en el nivel de complejidad sobre la máquina básica constituida por la cinesina y los microtubulos. La operación sinérgica de máquinas individuales como parte de un complejo estructural que funciona como una unidad y para un mismo propósito, nos permite hablar ya de un aparato.
Aparte de la organización radial y de la movilización coherente, existen otras dos importantes diferencias entre los cromatóforos y los demás sistemas de transporte intracelular examinados hasta ahora: 1) en todos los ejemplos anteriores el movimiento sobre los rieles del citoesqueleto es unidireccional o va en ambas direcciones, simultáneamente, pero no se alternan migraciones en un sentido y luego en otro; 2) el movimiento es continuo y no se presenta como respuesta ante factores extrínsecos a la propia célula. En los cromatóforos, por lo contrario, el pigmento tiene un comportamiento de ida y vuelta— se agrega o se dispersa en masa en un momento dado—, y la acción es inducida por hormonas o por impulsos nerviosos. En otras palabras, en los cromatóforos aparece un mecanismo de control. Esta particularidad es también propia de los aparatos.
Las evidencias acumuladas por la investigación de diversos aparatos de motilidad celular indican que la activación o inactivación de los mismos depende casi siempre de cambios en la abundancia de iones de calcio libres en el citoplasma. La mayoría de las células cuida celosamente que el calcio libre en su fluido interno se mantenga en un bajo nivel, al grado de que la concentración del mismo en el citoplasma es por lo regular 10 000 veces menor que en el medio extracelular. No obstante, ciertos estímulos pueden ocasionar que en algunas células la membrana plasmática permita la entrada de calcio, o que éste se escape de compartimientos internos donde es normalmente secuestrado con objeto de disminuir su presencia en libertad. Así, el aumento transitorio de calcio libre en el citoplasma tiene el efecto de apagar o encender el funcionamiento de las máquinas moleculares, o incluso de modificar la organización misma del citoesqueleto. En los cromatóforos la elevación del calcio libre en el citoplasma —a consecuencia de que hacen contacto con la membrana plasmática hormonas vertidas a la sangre del animal o sustancias secretadas por terminales nerviosas cercanas— parece obstaculizar la tracción centrífuga de la cinesina, ya que causa agregación del pigmento. Todavía no está esclarecida la secuencia de movimientos que determina esta respuesta. Sin embargo, es muy probable que tenga lugar una serie de procesos análogos a los que explican el control de la contracción de los músculos esqueléticos, y que tendremos oportunidad de revisar con detalle más adelante.
Migraciones de pigmentos retinales
Las migraciones masivas y reversibles de gránulos de pigmento no son exclusivas
de los cromatóforos. También existen en células de la retina de algunos animales,
donde cumplen con la función de regular la luz que llega a las células sensibles
o fotorreceptoras. Incluso se puede ver que en ciertos casos los fotorreceptores
cuentan con su dotación de pigmento transportable. Un buen ejemplo se encuentra
en los ojos de las moscas, mariposas y muchos otros insectos, y de crustáceos,
como los camarones y los cangrejos (Figura VI.3). Las partes propiamente fotosensibles
de las células fotorreceptoras pueden verse tapizadas por una densa cortina
de gránulos oscuros que evitan la iluminación excesiva de las mismas durante
las horas diurnas. Al anochecer, se retira el pigmento de esta posición y se
sitúa de tal manera que no impide el máximo aprovechamiento de la escasa luz
disponible.
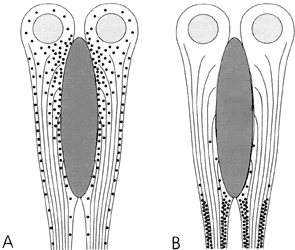
Figura VI. 3. Esquema ilustrativo de la distribución de los gránulos de
pigmento en los fotorreceptores retinales del cangrejo de río. A) En presencia
de la luz los gránulos se mueven hacia la parte superior para proteger las membranas
fotosensibles (zona alargada, gris oscuro); B) en la oscuridad los gránulos
se desplazan a la parte inferior, dejando descubiertas las membranas fotosensibles.
En los fotorreceptores del cangrejo de río este movimiento se efectúa por un
aparato de translocación en masa, integrado por un grueso haz de microtúbulos,
por una malla fibrosa equivalente a la red microtrabecular de los cromatóforos
y muy probablemente, por moléculas de cinesina. La estimulación lumínica origina
un aumento de calcio libre en el citoplasma de los fotorreceptores y dispara
el retorno de la población de gránulos de pigmento a la posición de adaptación
a la luz. Cuando entre los mecanismos que controlan el ingreso de la luz a la
retina se encuentran células pigmentarias —que no son fotosensibles —,
como en los peces y las ranas, el transporte se efectúa también por un citoesqueleto
especialmente organizado (Figura VI.4).

Figura VI. 4. Microscopía de barrido de alta resolución de un fotorreceptor
del cangrejo de río. Se pueden observar los gránulos de pigmento asociados a
haces de microtúbulos.
Hasta aquí hemos considerado movimientos celulares en donde los motores moleculares recorren grandes distancias sobre los rieles que proporciona el citoesqueleto. Cada motor está en libertad de desplazarse de uno a otro extremo del polímero respectivo. Tal vez la única excepción relativa en este sentido serían las migraciones pigmentarias en los cromatóforos y en las células retinales porque, a medida que alcanza su máxima extensión en cada región de la célula, la red microtrabecular del aparato de translocación impone probablemente cierta restricción a que continúe el avance de las moléculas de cinesina. En otros sistemas de motilidad, sin embargo, el recorrido de los motores sobre los rieles del citoesqueleto está limitado a distancias mucho menores. Paradójicamente, gracias a esta sujeción parcial se consiguen los más notables y poderosos movimientos biológicos. La actividad de los motores moleculares en el interior de las células es capaz, entonces, de realizar trabajo fuera de ellas, ya sea para acarrear materiales en el medio extracelular, o para transportar la célula completa de un lugar a otro, e incluso para movilizar al organismo entero del que forman parte. Todas estas proezas implican, por supuesto, aparatos muy complejos.
Cuando revisamos el caso del transporte axoplásmico se mencionó que el desplazamiento
en dirección al cuerpo celular parece estar mediado por la dinaína, un
translocador que migra sobre los microtúbulos en sentido opuesto al de la cinesina.
Sin embargo, la dinaína fue inicialmente conocida como el motor responsable
del movimiento de los cilios y los flagelos, unos apéndices filiformes de aproximadamente
0.2 micras de diámetro que emergen de la superficie de algunas células, y cuya
agitación al ritmo de entre 5 y 10 ciclos por segundo tiene una acción mecánica
sobre el líquido circundante. Cuando la célula se encuentra fija —como integrante
de un epitelio, por ejemplo— el batir de los cilios tiene un efecto de empuje
sobre el líquido externo, que es barrido sobre la superficie del tejido porque
muchas otras células idénticas realizan la misma operación. Un caso ilustrativo
de esto último es el epitelio que reviste las vías respiratorias, donde la función
de los cilios es encaminar al exterior el mucus que retiene las particulas
de polvo que penetran con el aire. Los óvulos de muchas especies también son
conducidos a través del oviducto, por medio de la actividad ciliar de células
epiteliales. Ahora bien, si se trata de una célula libre —como un protozoario
o un espermatozoide—, el trabajo de los cilios o los flagelos (Figura VI.5
y VI.7) se traduce en un desplazamiento de la propia célula a través del liquido
en que se encuentre, y constituye entonces un recurso de locomoción.
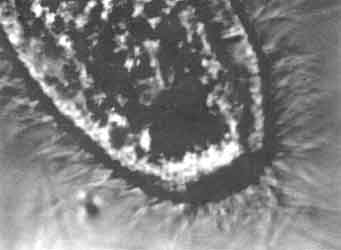
Figura VI. 5. Aspecto de los cilios alrededor de un paramecio vistos por
videomicroscopía de contraste, acentuada electrónicamente.
Para que los cilios y los flagelos puedan desempeñar su función con eficacia
necesariamente deben actuar con direccionalidad. Los cilios consiguen esto mediante
un movimiento análogo a las brázadas de un nadador: se extienden y empujan el
líquido como remos durante la fase activa del ciclo, y se repliegan hacia el
cuerpo de la célula en su retorno a la posición inicial (Figura VI .6) .
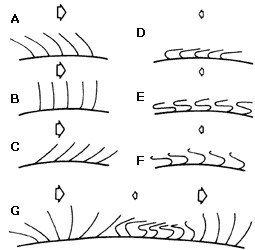
Figura VI. 6. Etapas del movimiento ciliar. A, B, C) Etapas de empuje; D,
E, F) etapas de retorno; G) cilios en etapas consecutivas del ciclo. Las flechas
indican magnitudes relativas y dirección de esfuerzos contra el medio.
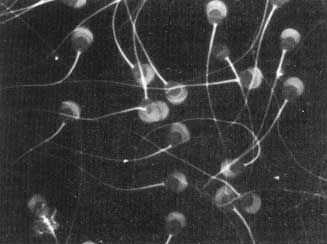
Figura VI. 7. Espermatozoides de cobayo en donde se ven claramente los flagelos
en diferentes etapas de reflexión. (Fotografía cortesía de A. Mújica).
Los flagelos —que son mucho más largos y generalmente menos numerosos que
los cilios— emplean un principio de propulsión similar al de una anguila:
ondulaciones continuas que recorren toda su longitud. Si el flagelo está inserto
en el polo posterior de la célula, como en los espermatozoides, la ondulación
dirigida hacia atrás representa un empuje hacia adelante. Cuando el flagelo
está situado en el polo anterior, como en muchos protozoarios y algas unicelulares,
el desarrollo de fuerzas es algo más complejo. El flagelo se curva hacia un
lado al enfrentar la resistencia del medio acuoso y realiza ondulaciones helicoidales
que obligan a la célula a girar con cierta inclinación en torno al eje de avance;
dicha rotación convierte al cuerpo celular en una especie de aspa que impulsa
el fluido hacia atrás y, en consecuencia, repercute en un desplazamiento del
organismo hacia adelante (Figura VI.8). Por increíble que parezca, esta diversidad
de movimientos es producto de un interdeslizamiento restringido de microtúbulos.
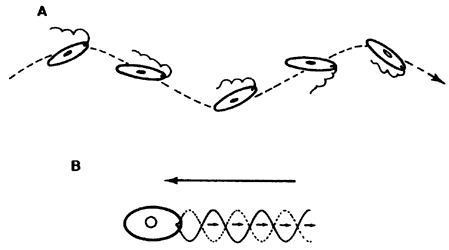
Figura VI.8. Representación esquematica de dos tipos de movimiento flagelar.
A) característica de algunos protozoarios y algas flageladas; B) característica
de espermatozoides. Las flechas grandes señalan la dirección de desplazamiento
del organismo.
La estructura básica de los cilios y los flagelos consiste en un aparato longitudinal al que los microscopistas llamaron axonema (filamento axial), recubierto por membrana plasmática como el resto del cuerpo celular (Figura III.9). Al igual que otros aparatos, el axonema está integrado por múltiples máquinas que trabajan de manera coordinada, y cada máquina consta de un motor y sus respectivos elementos de soporte. Estos últimos son provistos por un arreglo de microtúbulos paralelos y dispuestos de tal manera que un par central aparece circundado por nueve microtúbulos dobles. Los microtúbulos dobles periféricos no son simplemente parejas, como las que constituyen los dos centrales, sino unidades peculiares que resultan de una forma distinta de polimerización de la tubulina. Cada unidad está integrada por un microtúbulo incompleto, lo cual indica que no se cierra sobre sí mismo, adosado longitudinalmente a uno completo, de modo que la fracción faltante del primero queda sellada por la pared del segundo. Cada microtúbulo doble tiene su raíz en la corteza del cuerpo celular, donde se origina a partir de una estructura triple formada por la anexión lateral de un segmento más de microtúbulo incompleto; el conjunto resultante de nueve tripletes acomodados en formación anular recibe el nombre de cuerpo basal. Los dos microtúbulos centrales del axonema no llegan hasta el cuerpo basal ni presentan una inserción especial, pero se encuentran rodeados por una capa de material fibrilar llamada vaina interna.
Los microtúbulos dobles periféricos aparecen vinculados entre sí y con la vaina
interna por medio de varios tipos de enlaces perpendiculares al eje del axonema,
todos ellos colocados con exacta periodicidad a intervalos particulares. Desde
cada microtúbulo periférico se extienden puentes radiales hacia el centro del
axonema, y tenues bandas hacia uno de los dos microtúbulos dobles adyacentes.
Además, cada microtúbulo doble presenta dos hileras de brazos cortos que llegan
a tener contacto con el otro de los microtúbulos dobles vecinos. Estos brazos
son, las moléculas de dinaína o motores que, en combinación con los elementos
de apoyo estructural, constituyen las máquinas que mueven el axonema. El principio
del mecanismo no es difícil de comprender (Figura VI.9). Cuando dispone de
ATP, la dinaína propende a caminar sobre los microtúbulos en dirección
al cuerpo celular, arrastrando con ella cualquier carga unida, que en este caso
es también un mícrotúbulo doble con el que está permanentemente asociada. Sin
embargo, éste no puede migrar porque está anclado en el cuerpo basal y porque
se lo impiden además los múltiples vínculos con el centro del axonema y con
el microtúbulo doble anexo por el otro lado. En consecuencia, el esfuerzo produce
tan sólo un breve deslizamiento entre los dos microtúbulos conectados por la
dinaína. Dado que cada microtúbulo doble está sometido a un esfuerzo
similar, el resultado general es un efecto de torsión o flexión del axonema
en su conjunto.
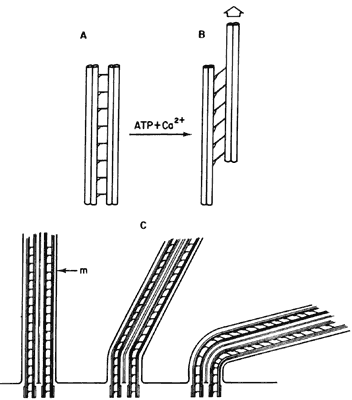
Figura VI.9. Mecanismo del axonema. Los pares exteriores están conectados
entre sí a través de otras proteínas y, al añadirse ATP y Ca2+,
el primero se hidroliza por la actividad de la dinaína para ceder la energía
necesaria para el movimiento. Las moléculas de dinaína asociadas a un par de
microtúbulos periféricos actúan sobre el par adyacente (A) y producen un esfuerzo
de deslizamiento en presencia de ATP y calcio (B). El esfuerzo
conjunto de todos los pares exteriores obliga a la flexión del cilio o flagelo
(C); m = membrana.
Por supuesto, nadie ha podido observar los vertiginosos fenómenos que ocurren
en el minúsculo interior de un cilio o un flagelo en movimiento. Pero existen
datos experimentales que sustenta la explicación resumida en el párrafo anterior.
No hay duda de que el mecanismo se encuentra en los cilios o flagelos mismos
porque, cuando son aislados de sus células— "rasurando'" protozoarios mediante
vibraciones de alta frecuencia o con ayuda de un fino rayo láser —, continúan
agitándose en una solución salina que contenga ATP. Es más, el
aparato motriz reside en el axonema porque la eliminación de la membrana plasmática
por métodos químicos no afecta la motilidad de cilios o flagelos aislados. Está
también claro que los motores son las moléculas de dinaína porque la exposición
del axonema a tratamientos capaces de retirarlas selectivamente causa un enlentecimiento
progresivo del movimiento, hasta llegar a la parálisis total, y la motilidad
se recupera al agregar dinaína purificada. Por otra parte, la aplicación de
enzimas que destruyen las conexiones pasivas entre los microtúbulos dobles sin
dañar mayormente los brazos de dinaína, seguida de adición de ATP,
da como resultado un rápido deslizamiento de dichos microtúbulos entre sí, hasta
que el axonema llega a medir nueve veces su longitud inicial (la suma de las
longitudes de los microtúbulos dobles individuales). Este fenómeno es una clara
indicación de que el movimiento ondulatorio de cilios y flagelos es consecuencia
de restricciones estructurales para que las moléculas de dinaína corran libremente
a lo largo de los microtúbulos con que hacen contacto funcional en el axonema.
Por último, el microscopio electrónico ha mostrado que los extremos de los microtúbulos
dobles en las puntas de los cilios, así como las diversas conexiones transversales
en la estructura del axonema, aparecen acomodados en posiciones que varían con
la curvatura del eje longitudinal, como se puede esperar según el modelo que
hemos descrito.
El axonema es una pequeña obra maestra de la ingeniería natural. Gracias a él los procesos internos del citoplasma logran influir mecánicamente sobre el medio exterior. Sin embargo, en este terreno los aparatos construidos con microtúbulos no pueden competir con aquellos que se basan en filamentos de actina. La potencia, velocidad y precisión de control que han alcanzado estos últimos les hacen acreedores al título de la maquinaria más perfecta y poderosa que ha creado la evolución. Es esta maquinaria molecular la que produce el brutal coletazo de una ballena, el raudo aleteo de un mosquito, el salto exacto de un trapecista y el arrebatador manejo del teclado de un virtuoso del piano. Su capacidad de transformación material del mundo ha quedado de manifiesto, con proporciones monumentales, en las pirámides de Egipto y Mesoamérica, las catedrales góticas de Europa y la Gran Muralla China. En cada uno de estos casos el movimiento implícito en la ejecución es o sido producto del trabajo muscular, y éste es la expresión macroscópica de la actividad de múltiples máquinas compuestas por moléculas de miosina y por filamentos de actína. Estas máquinas, en principio idénticas a las que generan las corrientes citoplásmicas en las plantas, conforman la unidad fundamental del músculo esquelético, un aparato contráctil conocido como sarcómero (del griego sarcos, carne, y meros, parte).
Los músculos esqueléticos —así llamados porque su trabajo ejerce fuerza
sobre los huesos en los vertebrados— están constituidos por muchas fibras
paralelas, cada una de las cuales se origina por la fusión de varias células
alargadas (Figura VI.1O(a). Puesto que al fundirse entre sí las células originales
eliminan las fronteras entre sus respectivos citoplasmas, pero conservan sus
núcleos individuales, las fibras musculares son en realidad grandes células
multinucleadas. Además, las fibras musculares presentan la singularidad de que
su citoplasma está surcado por numerosas fibrillas longitudinales formadas por
haces muy compactos de filamentos de actina y miosina. La cantidad de estas
proteínas en la fibra muscular es tan enorme que la mayor parte de su volumen
se encuentra ocupada por los polímeros correspondientes. Si bien las
fibras y las fibrillas pueden medir varios milimetros, y hasta centímetros de
largo, por lo común los filamentos individuales apenas exceden una micra de
longitud. Cada fibrilla está integrada por muchos segmentos de filamentos de
actina y miosina. El secreto del diseño del músculo esquelético, la clave de
su gran éxito evolutivo y de sus asombrosas capacidades radica en la perfecta
regularidad con que estos filamentos están distribuidos en el espacio (Figura
Vl.10(b).
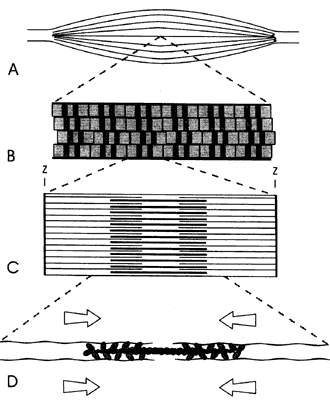
Figura VI.10. Organización de un músculo esquelético. A) El músculo está
formado por varias fibras; B) cada fibra muscular está formada por fibrillas,
constituidas, a su vez, por series de sarcómeros; C) el sarcómero esta formado
por filamentos de actina (delgados) y miosina (gruesos), que se interdigitan.
Los extremos de los filamentos de la actina se encuentran anclados en estructuras
terminales llamados discos Z; D) amplificación de la interacción que muestra
la colocación de las cabezas de la miosina en relación con los filamentos de
actina.
Cada fibrilla es una serie de miles de unidades discretas, y cada unidad está compuesta por un conjunto de filamentos de actina en cada extremo y un conjunto de filamentos de miosina en la región central. Los filamentos de actina de cada conjunto tienen la misma polaridad pero son antiparalelos con los del conjunto opuesto —tienen polaridad contraria—, y penetran parcialmente entre los filamentos de miosina. La polaridad de cada conjunto de filamentos de actina es tal que las moléculas de miosina caminan sobre ellos en el sentido de dirigirse hacia los extremos del sarcómero. Por su parte, los filamentos de miosina muestran la configuración bipolar que describimos en el capítulo anterior: las moléculas de miosina aparecen asociadas por sus colas, con las cabezas motoras colocadas de manera escalonada en dirección a los extremos. La unidad formada por estos tres conjuntos de filamentos es un sarcómero, y cada sarcómero está separado del que le sigue en la fibrilla por una placa circular llamada disco Z, en la que se anclan los filamentos de actina de ambos sarcómeros contiguos (Figura VI.10(c).
Con este tipo de ordenamiento la actividad motriz de las cabezas de miosina
tiene el efecto de tirar de ambos conjuntos de filamentos de actina y aproximarlos
entre sí. Esto es precisamente lo que sucede durante la contracción muscular;
los filamentos de actina son jalados por las cabezas de miosina. Como ocurre
con el deslizamiento de los microtúbulos dobles contiguos del axonema, la interacción
entre un filamento de miosina y un filamento de actina es mediada por múltiples
motores que operan de manera desfasada, por lo que en todo momento hay un cierto
número de ellos en la fase activa del ciclo mientras otros se hallan en las
etapas de reacomodo, quedando garantizada con ello una tracción sin interrupciones.
A diferencia del axonema, sin embargo, la organización del sarcómero brinda
la posibilidad de que la tracción sea aplicada de manera uniforme de cada filamento
y no sólo sobre uno de sus lados, como en los microtúbulos dobles de los cilios
y los flagelos. Esta uniformidad deriva de que los filamentos de miosina presentan
las cabezas motoras orientadas en varias direcciones en torno a su eje, por
lo que cada uno de ellos puede interaccionar con varios filamentos de actina
a la vez (Figura VI.10(d). Por su parte, cada filamento de actina hace contacto
con las cabezas motoras de todos los filamentos de miosina que lo circundan.
Puesto que dichas interacciones tienen lugar en ambos extremos de cada filamento
de miosina, los dos conjuntos de filamentos de actina experimentan un arrastre
uniforme y el sarcómero desarrolla una fuerza homogénea que tiende a disminuir
su longitud. Cada sarcómero tira entonces de los dos adyacentes y, dado que
todos los sarcómeros de una fibrilla y todas las fibrillas de una fibra se activan
prácticamente al mismo tiempo, el músculo entero se contrae (Figura VI. 11).

Figura VI.11. Micrografía electrónica mostrando el arreglo de los filamentos
de miosina y actina en los sarcómeros de músculo esquelético.
El grado de tensión y la amplitud del movimiento efectivo desarrollados por
los sarcómeros dependen de algunos factores. El más obvio es la carga que enfrenta
el músculo, porque si ésta es superior a la que puede vencerse con el empleo
de toda la potencia de los motores, la longitud de los sarcómeros no variará.
Esto es, lo que le sucede al novato que sin entrenamiento pretende levantar
una pesa de 140 kilos. El trabajo de las cabezas de miosina sobre los filamentos
de actina se realiza con un gran gasto de energía, pero la pesa permanece inmóvil
porque los músculos encuentran imposible acortarse ante una resistencia de tal
magnitud. El término que se emplea para designar este esfuerzo estático es contracción
isométrica —contracción sin cambio en las dimensiones del músculo —,
que puede parecer un contrasentido, pero en el fondo es válido, porque en realidad
algunos sarcómeros sí alcanzan a contraerse en cierta medida a expensas del
estiramiento de otros, y gracias a cierta elasticidad del tejido (Figura VI.12).
Ahora bien, no todas las contracciones isométricas son tan inútiles como la
del ejemplo anterior. Los músculos que mantienen a los animales de pie, contra
la fuerza gravitacional, cumplen con su función mediante contracciones isotérmicas
sostenidas. Cuando el animal se levanta a partir de una posición yacente, estos
músculos se contraen hasta que los huesos del esqueleto se encuentran con las
relaciones espaciales específicas que determinan la posición erguida para dicha
especie; una vez alcanzado este objetivo su actividad no puede ya expresarse
como cambio de longitud, el trabajo que desarrollan permite guardar dichas relaciones
y conservar la posición. Otro tanto ocurre al mantener un brazo alzado, al detener
una puerta que alguien empuja por el otro lado, y en cualquier otra situación
en la que oponerse a un movimiento debido a causas externas requiere de un esfuierzo
corporal continuo, aunque sea aparentemente estático.
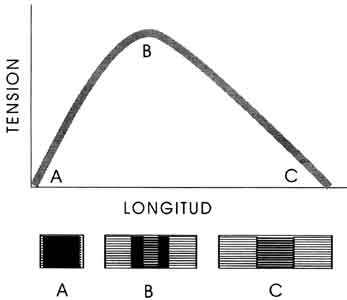
Figura VI. 12. Relación entre la longitud inicial de un sarcómero y la tensión
que puede desarrollar. La tensión es máxima en B cuando existe una interpenetración
media de los filamentos.
Por supuesto, mas que oponerse al cambio, la función principal del músculo es generar movimiento. Cuando se enfrenta con cargas proporcionales a su potencia particular, todo músculo activo se contrae efectivamente, es decir, reduce su longitud como resultado del acortamiento de sus sarcómeros. Sin embargo, debido a la construcción de estos últimos, la fuerza que pueden desarrollar al contraerse depende de su longitud inicial. Para comprender esta relación consideremos situaciones extremas. En un sarcómero excesivamente distendido antes de la contracción, la yuxtaposición de filamentos de actina y de miosina es mínima, puesto que el estiramiento aleja simétricamente a los primeros de la región media, donde se encuentran los segundos. En esta condición los dos grupos de filamentos se tocan entre sí sólo en sus porciones terminales y, por consiguiente, el número de cabezas de miosina que pueden interactuar con los filamentos de actina es reducido, y la fuerza total generada es también baja. Por lo contrario, en un sarcómero cuya longitud inicial sea demasiado corta, el traslapamiento de filamentos será máximo. En este caso, prácticamente la totalidad de las cabezas de miosina podrá ejercer tracción sobre los filamentos de actina pero, puesto que los filamentos de miosina se encontrarán en el tope de sus respectivos carriles —muy próximos a los discos Z—, el sarcómero se verá casi imposibilitado de contraerse más. De aquí que la posición más favorable para que un músculo desarrolle su máxima potencia es cuando sus sarcómeros tienen una longitud inicial intermedia, en forma tal que permita el contacto de numerosas cabezas de miosina con los filamentos de actina, y que a la vez ofrezca a los filamentos de miosina un buen trecho para desplazarse.
Esta relación entre el grado de estiramiento inicial y la potencia desarrollada
por el músculo esquelético no se presenta en otra gran clase de tejido contráctil:
el tipo de músculo que forma parte (de los vasos sanguíneos y de vísceras como
el estómago y los intestinos, la vejiga urinaria y el útero. Las paredes de
estos órganos pueden distenderse de manera considerable sin que disminuya su
capacidad de contracción, de la que depende su función de promover o regular
la transportación de los contenidos respectivos. Se dice, por lo tanto, que
esta clase de músculos tiene la propiedad de plasticidad , y la razón de esta
cualidad es estructural. Los filamentos de actina y miosina en las células de
este tejido no se encuentran dispuestos de manera especialmente ordenada, como
en el músculo esquelético. No hay en ellas series de aparatos unitarios equivalentes
a los sarcómeros, y por consiguiente no muestran las estiraciones periódicas
que caracterizan al músculo liso (Figura VI.13). La ventaja que significa la
plasticidad del músculo liso se ha conseguido, sin embargo, a costa de la rapidez
y precisión de trabajo que distinguen al músculo esquelético, pues los órganos
de que forma parte, no obstante que en ocasiones son muy poderosos, como es
el caso del útero, accionan con lentitud y sin una exacta coordinación espacio-temporal.
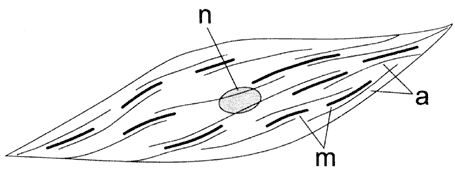
Figura VI. 13. Esquema de la organización de la miosina (m) y la actina
(a) en una célula de músculo liso. En estas células los filamentos no se arreglan
en forma de sarcómeros, sino que se distribuyen irregularmente en el citoplasma;
n = nucleo.
Una tercera categoría de músculo es la que constituye el corazón de los vertebrados. Este órgano excepcional, diseñado para funcionar sin interrupción durante toda la vida, posee atributos fisiológicos muy singulares. No obstante, desde el punto de vista de la organización de la máquinaria molecular que lo anima, es muy similar al músculo esquelético. Sus células son estriadas, puesto que los filamentos de actina y de miosina se encuentran dispuestos en forma de sarcómeros que se contraen de manera concertada y alternada entre diferentes regiones.
Hasta aquí hemos revisado las principales maquinarias moleculares y los aparatos que integran para efectuar la gran mayoría de los movimientos que presenta la materia viviente. Estos mecanismos comprenden a las máquinas constituidas por actina y miosina, o bien a las que tienen como base la tubulina y sus motores asociados. Estos dos sistemas operan simultáneamente en muchas células, cada uno encargado de su propia función. Pero al menos en la mayoría de las células animales llega un momento crucial en el que ambos conjuntos deben de actuar de manera perfectamente coordinada. Esta acción, una de las más asombrosas que haya producido la evolución, es la que consigue reproducir la vida misma —la división celular. La descripción de los aparatos que intervienen en este fenómeno fundamental es el tema del siguiente capítulo.